Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
ДНК-полимеразы могут находить и исправлять ошибки
Было установлено, что частота ошибок при репликации ДНК Е. coli не превышает 1 на 109-1010 нуклеотидов. Поскольку хромосома Е. coli содержит приблизительно 4,5 ∙ 106 пар оснований, на 10 000 клеток, претерпевших один цикл деления, встраивается всего один неправильный нуклеотид. Долгое время считалось, что столь высокая степень точности воспроизведения генетической информации целиком определяется точностью уотсон-криковского спаривания между матричной и новообразованной (дочерней) цепями, однако в результате последующего анализа выяснилось, что если бы точность репликации зависела исключительно от правильности спаривания оснований, то частота ошибок была бы значительно выше - приблизительно 1 на 104-105 остатков. Следовательно, чтобы объяснить такую низкую частоту ошибок при репликации in vivo, необходимо предположить участие в процессе репликации еще какого-то одного или нескольких факторов.
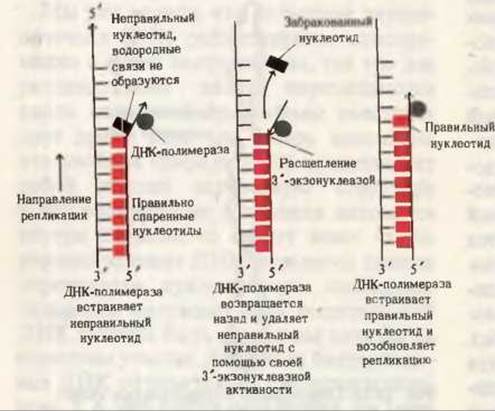
Рис. 28-15. Исправление ошибок с помощью 3'-экзонуклеазной активности ДНК-полимеразы.
Более детальное изучение свойств высокоочищенных ДНК-полимераз позволило получить по крайней мере частичный ответ на вопрос о природе этих факторов. Напомним, что ДНК-полимеразы I и III обладают тремя различными ферментативными активностями. Мы уже видели, как фермент функционирует в качестве полимеразы, а также как он может удалять нуклеотидные остатки с 5'-конца фрагмента ДНК. Однако 3'-экзонуклеазная активность ДНК-полимераз I и III очень озадачивала исследователей, ибо она означала, что эти ферменты способны “пятиться”, отщепляя 3'-концевые нуклеотиды в направлении, противоположном тому, в котором они действуют как полимеразы. 3'-экзону- клеазная активность ДНК-полимераз I и III - это средство проверки новосинтезированной цепи ДНК и исправления ошибок, сделанных ферментом при его работе в качестве полимеразы. Если ДНК-полимераза встраивает неправильный нуклеотид, то фермент сам может распознать неспособность этого нуклеотида образовать правильную пару с соответствующим нуклеотидом матрицы (рис. 28-15). В этом случае фермент возвращается назад и отщепляет неправильный нуклеотид с 3'-конца цепи, после чего полимераза продолжает присоединять правильные нуклеотиды, т.е. возобновляет свое обычное продвижение в направлении 5'→3'. Таким образом, по мере перемещения репликативной вилки вдоль матрицы осуществляется проверка каждого встроенного нуклеотида. Корректирующее действие ДНК-полимеразы очень эффективно; благодаря ему точность репликации повышается как минимум в 104 раз. Суммарная ошибка возникает в результате ошибок, допускаемых ферментом в ходе полимеризации и в процессе исправления их при корректировке; она не превышает одной ошибки на 109-1010 нуклеотидных остатков.
Очень важно отметить, что процесс репликации протекает со значительно более высокой степенью точности, чем процессы транскрипции и трансляции. Частые ошибки в репликации подвергли бы большому риску сохранность видов и их жизнеспособность. Ошибки же в транскрипции и трансляции гораздо менее опасны, поскольку они влияют на образование РНК или белка только в одной клетке и не изменяют всю последующую родословную вида. Корректировка с помощью ДНК-полимеразы - это, вероятно, лишь один из путей, обеспечивающих высокую точность репликации. Возможно, исключительно сложная организация репликативного процесса и участие в нем множества белков необходимы для достижения именно этой цели. Интересно, что некоторые эукариотические ДНК-полимеразы не осуществляют корректировку. По-видимому, эукариоты обеспечивают точность и надежность процесса репликации с помощью каких-то других средств.