Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Еще о генах: репарация, мутации, рекомбинация и клонирование
Разнообразие антител - это результат перемещений и рекомбинаций
Один из наиболее необычных типов генетической рекомбинации наблюдается в процессе образования генов, которые определяют синтез различных антител у позвоночных. Антитела или иммуноглобулины - это белки, продуцируемые лимфоцитами (иммуноцитами) позвоночных в ответ на вторжение в организм чужеродной макромолекулы - антигена (разд. 6.11). Каждый тип антигена способен связываться с иммуноцитами одного специфического типа, вызывая рост и деление этих клеток и приводя к образованию уникального клона, т.е. линии полностью идентичных иммуноцитов. Клетки такого клона вырабатывают только один вид иммуноглобулинов, специфически связывающих только тот антиген, который индуцировал их размножение. В результате взаимодействия антигена с антителом образуется комплекс, в котором антиген обычно утрачивает свою биологическую активность. Удивительным является тот факт, что человеческий организм умеет синтезировать буквально миллионы разных видов антител, каждый из которых способен связываться только с одним из миллионов различных антигенов, воздействию которых организм может подвергнуться. Антитела образуются не только против большинства белков других животных, бактерий, вирусов, паразитов и растений, но практически против любой макромолекулы, даже искусственного происхождения.
Может создаться впечатление, что для того, чтобы иметь возможность синтезировать различные антитела, каждое из которых взаимодействует лишь с одним из многих миллионов возможных антигенов, организм должен содержать соответствующее число разных генов для каждого из возможных антител в отдельности. Долгое время это казалось невероятным. В ядрах клеток человека просто нет столько ДНК, чтобы кодировать миллионы различных антител в дополнение ко многим тысячам обычных белков, определяющих структуру, метаболизм и индивидуальность человеческого организма.
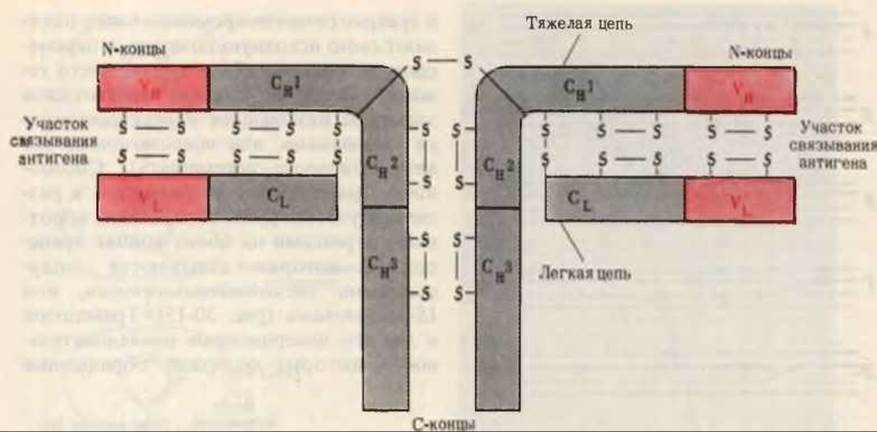
Рис. 30-16. Структура молекулы антитела. Молекула содержит две легкие (L) и две тяжелые (Н) цепи, каждая из которых имеет вариабельную (V) (красная) и константную (С) области (серая). Константные области тяжелых цепей состоят из трех различающихся доменов Сн1, Сн2 и Сн3. Молекула содержит большое число дисульфидных мостиков и углеводных остатков, прикрепленных к тяжелым цепям.
Ответ на эту загадку был получен в результате исследования структуры антител и генов, которые их кодируют. На рис. 30-16 показана структура антитела, мол. масса которого составляет приблизительно 160000. Молекула антитела состоит из двух тяжелых, т. е. длинных, полипептидных цепей (по 446 аминокислотных остатков в каждой) и из двух легких, т.е. коротких, цепей (по 214 аминокислотных остатков). Цепи соединены между собой поперечными дисульфидными мостиками; кроме того, в обеих цепях имеются еще и внутренние-S—S-связи. Каждая тяжелая и каждая легкая цепь содержит область с неизменной аминокислотной последовательностью, характерной для данного вида; она называется константной, или С-областью. В каждой цепи находится также вариабельная V-область, аминокислотная последовательность которой, по-видимому, различна у антител разных типов. Константная область тяжелых цепей состоит из трех доменов, имеющих сходные аминокислотные последовательности. Молекула антитела содержит два антигенсвязывающих участка, каждый из которых расположен в углублении (“кармане”) между концами вариабельных областей тяжелой и легкой цепей. Молекулярная модель антитела представлена на рис. 30-17.
Наличие в антителах константных и вариабельных областей уже давно навело ученых на мысль, что ДНК, направляющая синтез легких цепей, образуется в результате сплайсинга двух генов, один из которых кодирует вариабельную область, а другой - константную. Доказательства, подтверждающие это предположение, были получены Сусуму Тонегавой и его сотрудниками, которые показали, что гены, кодирующие константную и вариабельную части легких цепей одного специфического типа, сильно сближены в ДНК иммуноцитов, вырабатывающих данный тип молекул, тогда как в ДНК другой линии иммуноцитов, не способной продуцировать этот тип легких цепей, они расположены далеко друг от друга. Тонегава и сотр. сделали вывод, что иммуноциты могут выбирать участки ДНК, кодирующие вариабельные части молекулы антитела, из разных и далеко отстоящих друг от друга областей генома клетки и перемещать их в место, расположенное следом за геном, кодирующим константную часть данного типа легких цепей. После соединения генов в одно целое РНК-полимераза может обеспечить синтез единой молекулы мРНК, кодирующей всю легкую цепь.
В результате дальнейших исследований выяснилось, что ДНК, кодирующая вариабельные области как легких, так и тяжелых цепей, состоит из генов нескольких типов, которые могут менять свое положение и собираться вместе с образованием разнообразных комбинаций (рис. 30-18). ДНК, определяющая вариабельные области антител, составлена приблизительно из четырехсот различных вариабельных (V) генов, двенадцати так называемых генов разнообразия (D-гены) и четырех соединительных (J) генов. Сочетание этих генов в различных комбинациях позволяет составить ДНК более чем для 20 000 или даже большего числа вариабельных областей. Эти гены в свою очередь претерпевают дополнительные изменения в своих нуклеотидных последовательностях и присоединяются к различным участкам ДНК, кодирующим константные области, в результате чего образуются миллионы разных генов, ответственных за синтез антител. Многие вопросы синтеза антител ждут еще своего решения, однако уже сейчас ясно, что перемещение и рекомбинация генов или частей генов - это очень эффективный и точно воспроизводящийся процесс, при помощи которого иммуноциты способны вырабатывать антитела против практически любой макромолекулы.
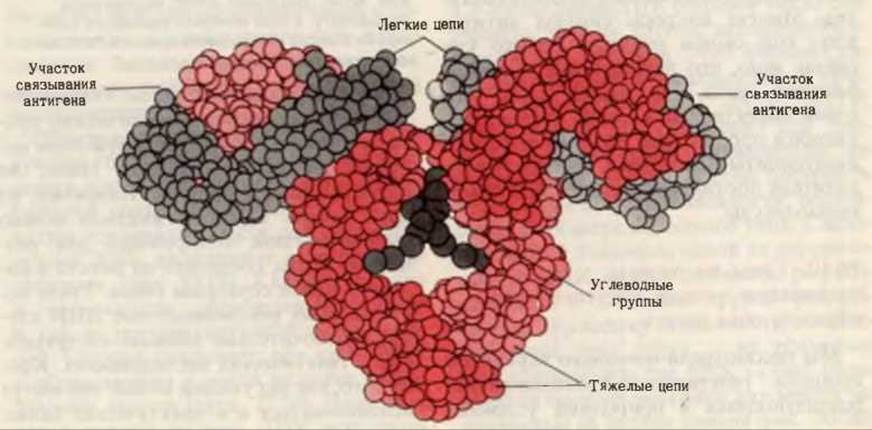
Рис. 30-17. Модель структуры иммуноглобулина, построенная на основании данных рентгеноструктурного анализа.