БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 9.НУКЛЕЇНОВІ КИСЛОТИ
9.5. Структура й функції рибонуклеїнових кислот
9.5.3.Структурна організація РНК
У всіх прокаріотичних і еукаріотичних організмів є три основні типи молекул РНК (мРНК, рРНК, тРНК), які розрізняють за первинною структурою, молекулярною масою, конфігурацією, тривалістю життя і, найголовніше, за функціональною активністю.
Рибосомні РНК. Частка рРНК становить близько 80 % всієї РНК клітини. У клітинах E. coli виявлено три типи рРНК: 5S, 16S і 23S (S-позасистемна одиниця (сведберг), яка має розмірність часу (1S = 10-13 с) і характеризує молекулярну масу, об'єм і форму макромолекул)), а в еукаріотичних клітинах функціонують чотири типи - 28S, 18S, 5,8S і 5S рРНК.
Рибосомні РНК характеризуються вищим, ніж звичайно, вмістом гуаніну й цитозину. Зустрічаються також декілька модифікованих нуклеотидів (приблизно 1 %), які в переважній більшості метильовані за азотистою основою або рибозою (2'-метилрибоза).
Вторинна структура рРНК представлена значною кількістю коротких двоспіральних ділянок і петель. На рис. 9.17 зображено структури 16S рРНК клітин E. coli та дріжджів. Незважаючи на те, що нуклеотидна послідовність у них різна, вторинні рівні організації молекул дуже схожі.
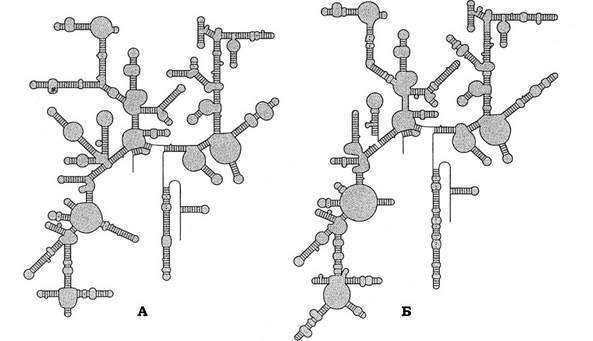
Рис. 9.17. Вторинний рівень організації 16S рРНК клітин E. coli (А) і Saccharomyces cervisiae (дріжджі) (Б)
Транспортні РНК - становлять близько 15 % усієї РНК клітини. Молекули тРНК складаються зазвичай із 75-85 нуклеотидів. Молекулярна маса тРНК 25-28 кД (4S). Кількість різних тРНК - декілька десятків: від 1 до 6 видів для кожної із 20 протеїногенних амінокислот. Види тРНК, які здатні зв'язувати одну й ту ж амінокислоту, називаються ізоакцепторними.
Для нуклеотидного складу тРНК характерним є переважання гуаніну й цитозину (60 %о), а також досить високий вміст мінорних, або модифікованих, нуклеотидів (8-19 %). До їхнього складу відносять дигідроуридин, псевдоуридин, інозин, метилінозин, метил- і диметилгуанозин, метилуридин (риботимідин). Частка спарених основ дещо перевищує 75 %, причому лише частина з них (до 60 %) припадає на спіралізовані ділянки, а решта водневих звязків замкнуті між основами з різних неспіралізованих ділянок полінуклеотидного ланцюга.
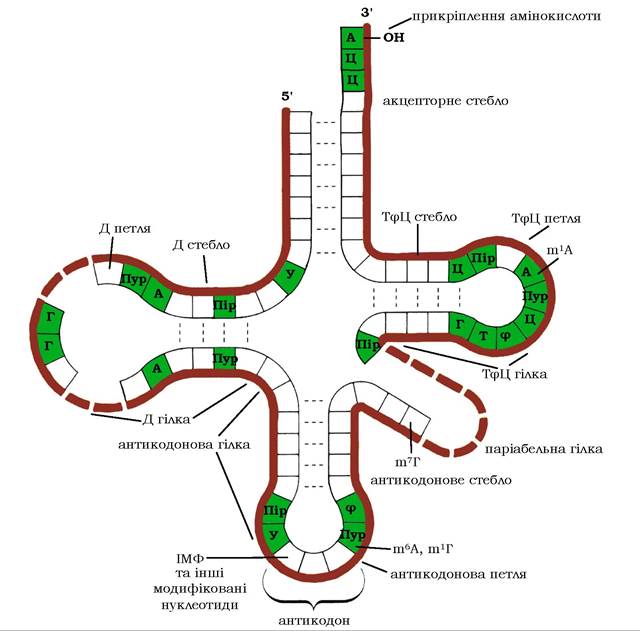
Рис. 9.18. Схема вторинної структури молекули тРНК.
Вторинна структура молекул тРНК у двовимірному просторі має конформацію "листка конюшини» (рис. 9.18). У цій структурі - чотири дволанцюгові та п'ять одналанцюгових ділянок. Оскільки мінорні нуклеотиди, як правило, не здатні до комплементарної взаємодії, вони містяться переважно в одноланцюгових ділянках. Неспарені нуклеотидні послідовності формують такі специфічні для будови тРНК структурні елементи:
✵ акцепторне стебло (гілка) складається з чотирьох лінійно розташованих нуклеотидів, три з яких мають в усіх видів тРНК однакову послідовність - ЦЦА. Кінцевий аденозин через 3'-ОН- групу рибози акцептує амінокислоту в процесі синтезу білка.
✵ антикодонове стебло (гілка) містить ділянку із 7 нуклеотидів у середній частині ланцюга, три з яких виконує функцію антикодону, який комплементарно взаємодіє з відповідним кодоном у ланцюгу мРНК.
✵ дигідроуридилове стебло (Д-петля) - складається зі стебла в 3-4 пари нуклеотидів і петлі варіюючого розміру: 8-12 нуклеотидів, до складу яких входить 1-4 дигідроуридилові залишки. Припускають, що Д-петля необхідна для зв'язування з ферментом аміноацил-тРНК-синтетазою, який бере участь у впізнаванні амінокислотою своєї тРНК.
✵ псевдоуридилове стебло (ΤΨЦ-петля) містить шпильку із 7 нуклеотидів, яка складається з двох незвичайних залишків: рибо- тимідин (Т) і псевдоуридин (Ψ). Тринуклеотид ТΨЦ завжди розташований в одному й тому ж місці петлі. Вважають, що ТΨЦ-петля бере участь у зв'язуванні тРНК з рибосомою.
✵ варіабельна гілка являє собою структуру, за кількістю нуклеотидних залишків у якій тРНК поділяються на два класи: тРНК класу I (75 % від загальної кількості) містять додаткову гілку довжиною 3-5 нуклеотидів. Додаткова гілка тРНК класу II складається із 13-21 нуклеотидів і часто включає неспарену петлю.
Молекули різних тРНК відрізняються одна від одної послідовністю нуклеотидів, проте їхня третинна структура схожа. Молекула має такий характер упакування, що чотири дволанцюгові ділянки попарно зближені й утворюють приблизно два витки подвійної спіралі, розташовані майже перпендикулярно один до одного, і молекула набуває Г-подібної форми (рис. 9.19).
Матричні РНК - найбільш гетерогенні щодо розмірів (константа седиментації від 6 до 25S) і стабільності молекули, їхня частка від загальної маси РНК у клітині невелика - близько 5 %.
Матричні РНК, особливо еукаріотичні, мають деякі унікальні структурні особливості. Так, 5'-кінець кепований метилгуанозин- трифосфатом, приєднаним до 5'-гідроксилу сусіднього 2'-О-метилрибонуклеозиду через залишок трифосфату (рис. 9.20). Молекули мРНК часто містять внутрішні залишки 6-метилгуаніну та 2'-О-метильовані рибонуклеотиди. Вважають, що кепування мРНК з 5'-кінця використовується для специфічного впізнавання в системі трансляції (на стадії ініціації), сприяє стабілізації молекули, захищаючи її 5'-кінці від дії фосфатаз і 5'-екзонуклеаз, а також бере участь у реакціях процесингу молекули мРНК.
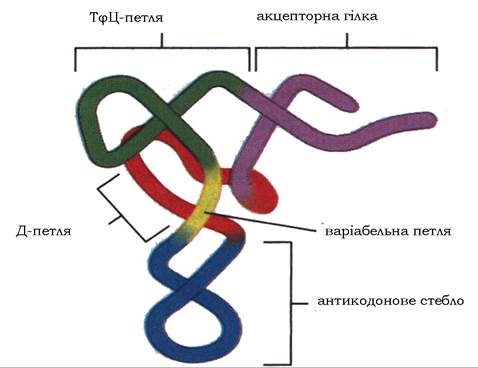
Рис. 9.19. Схема третинної структури молекули тРНК
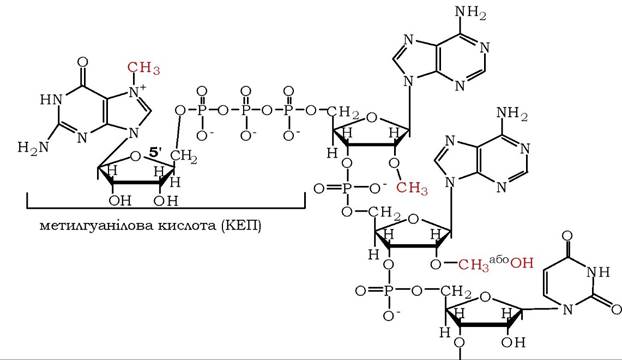
Рис. 9.20. Кеп-структура на 5'-кінці більшості еукаріотичних мРНК
За кеп-структурою розташована 5'-нетранслююча ділянка - послідовність з декількох десятків нуклеотидів, яка комплементарна одній із ділянок рРНК, що входить до складу малої субодиниці рибосоми. За рахунок цього мРНК первинно зв'язується з рибосомою на підготовчій стадії трансляції.
Протилежний кінець (З'-кінець) більшості молекул мРНК містить поліаденілатний ланцюг із 50-250 нуклеотидів. Вважають, що наявність подібної структури (полі (А) - "хвіст") сприяє підтримуванню внутрішньоклітинної стабільності мРНК. Можливо також, що поліаденілювання забезпечує транспорт мРНК у цитоплазматичне середовище.