БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 10. МЕТАБОЛІЗМ НУКЛЕЇНОВИХ КИСЛОТ
10.6.Біосинтез ДНК (реплікація)
10.6.2. Механізм реплікації ДНК
Біосинтез ДНК складається з великої кількості послідовних етапів, які включають упізнавання точки початку реплікації, розплетення подвійної спіралі батьківської молекули, утворення репліка- тивної вилки (ініціація), синтез нових ланцюгів (елонгація) і завершення синтезу двох дочірніх ланцюгів ДНК (термінація).
Ініціація реплікації. Оскільки синтез ДНК відбувається на одноланцюговій матриці, йому має передувати обов'язкове розділення двох ланцюгів ДНК. Процес починається в певній точці (ориджин) одночасно в обох напрямках через утворення реплікативних вилок. У цих точках реплікації на ДНК діють особливі білки - ДНК-топоізомерази, які вносять у ланцюг ДНК одно- або дволанцюгові розриви з наступним скиданням витків спіралі та
відновленням безперервності ланцюгів. Сполучення розірваного ланцюга відбувається без додаткових енергетичних витрат, оскільки необхідна енергія запасається у вигляді макроергічного ковалентного зв'язку, який виникає між пентозофосфатним каркасом ланцюга ДНК і топоізомеразою. За такої дії ДНК-топоізомераз розділення двох ланцюгів у реплікативній вилці відбувається легше. Розрив водневих зв'язків, розходження полінуклеотидних ланцюгів і просування реплікативної вилки забезпечується ферментами ДНК-хеліказами, які для своєї роботи потребують енергії гідролізу АТФ. Далі з одноланцюговими ділянками ДНК зв'язуються спеціальні спіральдестабілізуючі білки - SSB-білки, які не дозволяють одиночним ланцюгам ДНК замкнутися (ренатурувати). Подібна вибіркова спорідненість SSB-білків до одноланцюгових ділянок ДНК пояснюється наявністю в них гідрофобних зон, куди проникають азотисті основи. Крім того, SSB-білки кооперативно взаємодіють один з одним, вишиковуючись у довгі ряди вздовж одноланцюгової ДНК і таким чином стабілізують її в такому стані.
Увесь комплекс білків, які забезпечують процес реплікації, утворює в реплікативній вилці ДНК складну надмолекулярну структуру, яку назвали реплісомою.
Оскільки ДНК-полімерази не здатні самостійно почати синтез ланцюга, то необхідний інший фермент, який забезпечує праймерний (приманковий) полінуклеотидний ланцюг. Цей фермент, ДНК-праймаза, не потребує для ініціації синтезу вільної 3'-ОН-групи, копіює частину матричного ланцюга ДНК у напрямку від 5'- до З'-кінця і в такий спосіб утворює коротку (10-60 рибонуклеотидів) одноланцюгову РНК (РНК-приманку) з 3'-ОН вільним кінцем. Далі ДНК-праймаза вивільнює місце, а ДНК-полімераза використовує 3'-ОН групу приманки для продовження процесу полімеризації ланцюга ДНК на стадії елонгації реплікації.
Елонгація реплікації. Синтез нових ланцюгів ДНК здійснюється з урахуванням принципу комплементарності: кожний нуклеотид, що підбирається в зростаючий (дочірній) ланцюг, повинен бути комплементарним відповідному нуклеотиду в матричному (батьківському) ланцюзі.
ДНК-полімерази здійснюють процес полімеризації нуклеотидів тільки в напрямку 5'→3', а переміщення ферменту відбувається в напрямку 3'^5' матриці. Виходячи з цього, безперервне продовження елонгації зростаючого ланцюга може відбуватися лише вздовж одного ланцюга-матриці, того, відносно якого реплікативна вилка рухається від 3'- до 5'-кінця. Ланцюг, що синтезується безперервно, називається лідируючим (рис. 10.17).
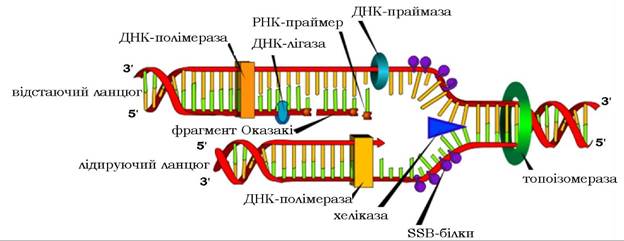
Рис. 10.17. Схема реплікації ДНК
На другому ланцюзі-матриці синтез нового ланцюга здійснюється в напрямку 5'→3', але проти руху реплікативної вилки. Тому другий ланцюг синтезується уривчасто, короткими фрагментами, які називають фрагментами Оказакі (за прізвищем дослідника, що їх відкрив), і має назву відстаючого.
Кожний фрагмент Оказакі починається з відповідного РНК-праймера й закінчується перед початком передуючого йому праймера. У прокаріотів довжина фрагмента Оказакі становить близько 1000, а в еукаріотів 100-200 нуклеотидів. Така різниця в довжині фрагментів Оказакі та різниця у швидкості синтезу ДНК пояснюється наступним. На відміну від прокаріотів, реплікація ДНК в еукаріотів відбувається не у вільному стані, а у складі хроматину. А, як відомо, хроматин складається з нуклеосом, на утворення яких витрачається по 200 пар нуклеотидів ДНК. Можливо, що саме тому в еукаріотів фрагменти Оказакі відстаючого ланцюга ДНК мають довжину 100-200 нуклеотидів. За сучасними уявленнями, існують нуклеосоми, які створюють бар'єр, здатний затримувати рух ДНК-полімерази. Отже, стає зрозумілим той факт, що швидкість реплікації в еукаріотів значно менша за таку в прокаріотів.
ДНК-полімераза ІІІ прокаріотів (ДНК-полімераза δ еукаріотів) послідовно нарощує ланцюг, один за одним приєднуючи до нього відповідні дезоксирибонуклеотиди (рис. 10.18). Однією ділянкою активного центру фермент захоплює перший неспарений нуклеотид матриці в зоні реплікативної вилки, другою - зростаючий кінець полінуклеотидного ланцюга, що містить вільну ОН-групу на 3'-кінці, третьою зв'язує один із вільних дезоксирибонуклеозидтрифосфатів, який комплементарний неспареному нуклеотиду матриці та розташований у першій ділянці активного центру. У такому стані фермент атакує α-фосфодіефірний зв'язок зв'язаного нуклеозидфосфату, при цьому відщеплюється пірофосфат, а утворений нуклеозидмонофосфат 3',5'-фосфодіефірним зв'язком приєднується до 3'-ОН-групи зростаючого ланцюга.
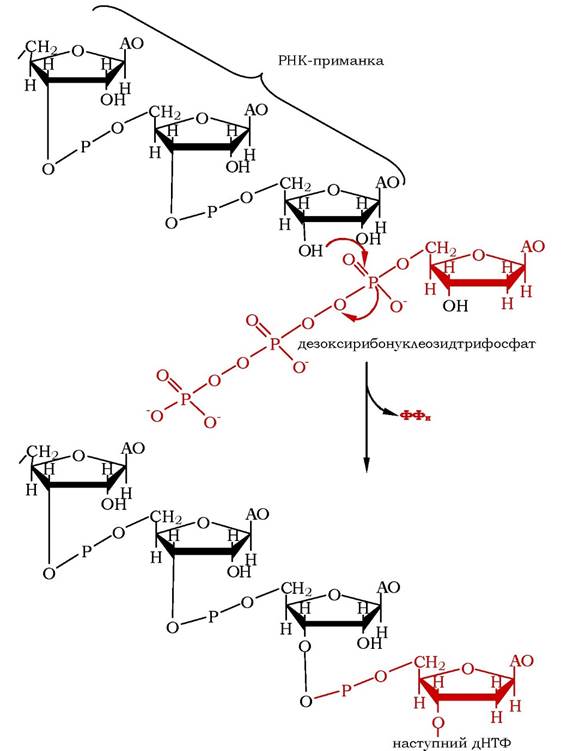
Рис. 10.18. Елонгація реплікації ДНК
Використання дНТФ робить процес утворення фосфодіефірних зв'язків термодинамічно вигідним, оскільки утворення фосфодіефіру з монофосфату й ОН-групи відбувається спряжено з гідролізом ангідридного пірофосфатного зв'язку.
Крім полімеризації ланцюгів під час реплікації ДНК відбувається вирізання РНК-праймерів з лідируючого ланцюга та з кожного фрагмента Оказакі. Цю функцію в прокаріотів виконує ДНК-полімераза І (аналогічну функцію в еукаріотів виконує ДНК-полімераза β). Фермент поступово видаляє з 5'-кінця праймера по одному рибонуклеотиду за допомогою своєї 5'→3'-екзонуклеазної активності. У цей час до ОН-групи на З'-кінці попереднього фрагмента Оказакі ДНК-полімераза приєднує дезоксирибонуклеотиди в кількості, яка дорівнює вирізаному праймеру, і таким чином заповнює прогалину, що виникла при видаленні рибонуклеотидів.
Сполучення фрагментів ДНК на відстаючому ланцюзі відбувається за допомогою ферменту ДНК-лігази, яка каталізує утворення фосфодіефірного зв'язку між 5'-фосфатом одного фрагмента ланцюга і 3'-ОН групою дезоксирибози наступного фрагмента з утворенням безперервного ланцюга ДНК.
На відміну від полімеразної реакції за участю ДНК-полімерази, де одним з учасників процесу є вільний дезоксирибонуклеозидтрифосфат, у ДНК-лігазній реакції обидва учасники - кінцеві дезоксирибонуклеозидмонофосфати містяться в складі фрагментів, що об'єднуються. Крім того, ДНК-лігази сполучають тільки такі одноланцюгові фрагменти, які знаходяться у складі дволанцюгової ДНК.
Термінація реплікації. Термінація реплікації ДНК залежить від структури ДНК. Якщо відбувається реплікація кільцевої ДНК безперервний ріст лідируючого та відстаючого ланцюгів уздовж матриці неминуче приводить до зміщення 3'-ОН- та 5'-фос- форильних кінців одного ланцюга в точці початку реплікації. При двонаправленій реплікації кільцевої ДНК, коли утворюються дві реплікативні вилки, що рухаються у двох протилежних напрямках, реплікація ДНК завершується на відстані 180° від точки початку синтезу. Кільцеві ДНК у місцях зустрічі сполучаються ДНК-лігазою, при цьому вони виявляються попарно зчепленими. Надалі відбувається їхнє розділення на окремі геноми за допомогою топоізомерази типу ІІ.
У випадку термінації та завершення реплікації лінійних ДНК процес відбувається шляхом розділення геному на численні окремі реплікони. Такі реплікони активуються не всі одночасно, проте (як у еукаріотів) клітинному поділу має передувати обов'язкова одноразова реплікація кожного з них. Зупинка просування реплікативної вилки відбувається тільки за умови зіткнення з іншою вилкою, яка рухається в протилежному напрямку, або при досягненні кінця хромосоми. Унаслідок цього вся ДНК хромосоми за короткий час стає реплікованою.
На завершення реплікації утворюються два ланцюги ДНК, ідентичні початковим її ланцюгам. Кожний із батьківських ланцюгів залишається інтактним і служить матрицею для синтезу комплементарного ланцюга. Утворений таким чином новий подвійний ланцюг містить один вихідний, а інший - синтезований.
Реплікація теломерних ділянок ДНК. Кінцеві області еукаріотичних хромосом - теломери - містять специфічну нуклеотидну послідовність, представлену численними тандемно розташованими олігонуклеотидними (гексануклеотиди - АГГГТТ) послідовностями. Така послідовність називається теломерною (теломерна ДНК). Загальна довжина таких повторів на одному кінці ДНК на початковій стадії онтогенетичного розвитку організму становить від 10 до 15 тисяч нуклеотидних пар.
Наявність теломерних повторів необхідна для завершення реплікацій кінцевих інформативних послідовностей у ДНК хромосом, тобто для збереження генетичної інформації.
Після завершення реплікації лінійної молекули ДНК 5'-кінці її дочірніх ланцюгів залишаються недореплікованими, оскільки на цьому кінці містяться РНК-приманки, які були видалені (рис. 10.19).
Важливо зазначити, що у випадку реплікації кільцевих бактеріальних ДНК цієї проблеми не виникає, оскільки перші утворені РНК-приманки видаляються ферментом, який одночасно заповнює утворену прогалину шляхом нарощування 3'-ОН кінця зростаючого ланцюга ДНК.
Теломерні повтори не несуть генетичної інформації, тому втрата деякої частини цих послідовностей не приводить до порушення функціонування геному - своїм існуванням вони захищають від недореплікації важливіші послідовності ДНК.
З кожним клітинним поділом ДНК хромосом вкорочується, що становить близько 0,00005 % її довжини. На перший погляд, це досить незначна величина і нею можна було б нехтувати. Але якщо врахувати, що кількість клітинних поділів велика, то відсутність механізмів відновлення довжини теломерних ділянок ДНК хромосом може привести до катастрофічних наслідків.
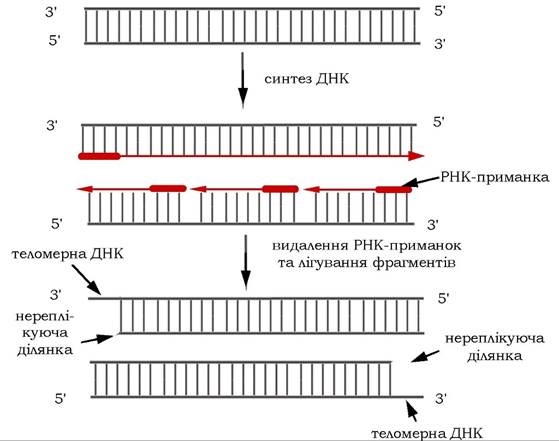
Рис. 10.19. Укорочення заново синтезованих ланцюгів ДНК після видалення РНК-приманок
Кінцева недореплікація ДНК хромосом має велике біологічне значення. За існуючими уявленнями, укорочення теломер до критичного рівня веде до початку старіння соматичних клітин, а при досягненні цього рівня - до їхньої смерті (теорія теломерного старіння).
Проте в зародкових, стовбурових та інших клітинах, що швидко діляться, існує механізм, за якого добудовуються недорепліковані ділянки ДНК. В еукаріотичних клітинах існує фермент термінальна дезоксинуклеотидилтрансфераза (теломераза), яка забезпечує відновлення недореплікованих 5'-кінців. Як простетичну групу теломераза містить РНК довжиною близько 450 пар нуклеотидів. Вона одночасно служить матрицею при синтезі теломерних повторів і для зв'язування шляхом гібридизації з кінцевою теломерною ділянкою ДНК. За допомогою РНК теломераза за принципом комплементарності подовжує не дочірній, а батьківський ланцюг у напрямку 5'→3' (рис. 10.20).
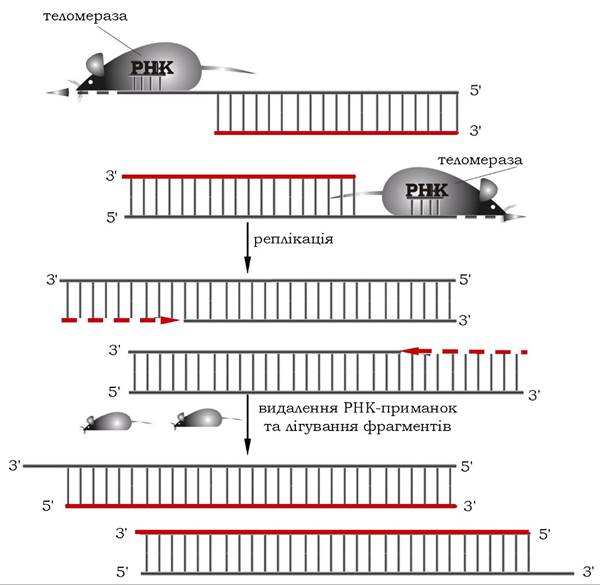
Рис. 10.20. Подовження теломерної ділянки ДНК за участю теломерази
До 3'-кінця ланцюга ДНК теломераза послідовно приєднує гексануклеотид АГГГТТ. Потім вона зміщується по ланцюгу ДНК на один теломер і починає синтез нового гексануклеотиду. Нарощення ланцюга за допомогою теломерази відбувається доти, поки подовжений ланцюг здатен виступати як матричний для утворення ще одного фрагмента Оказакі нового ланцюга.
Спочатку до дочірнього ланцюга праймаза синтезує РНК- приманку, а потім ДНК-полімераза β послідовно приєднує до приманки дезоксирибонуклеотиди, що комплементарні теломерним повторам батьківського ланцюга. Реплікація фрагмента відбувається в напрямку 5'→3' і припиняється при зіткненні з попереднім 5'-кінцем дочірнього ланцюга. Сполучення (лігування) фрагмента з ланцюгом здійснюється за допомогою ДНК-лігази, а екзонуклеаза видаляє РНК-затравку з 5'-кінця нарощеного дочірнього ланцюга.
Таким чином кінець дволанцюгової ДНК набуває вихідної конформації, яка була до дії теломерази, хоча дочірний ланцюг є коротшим за батьківський, але фактично довшим на серію теломерних повторів.
У більшості соматичних клітин організму механізм реплікації теломер відсутній - теломераза неактивна й тому при поділі клітин теломерна ДНК поступово скорочується. Такі клітини мають довжину теломерної ДНК достатню для часу життя клітини та її нащадків. Проте відомо, що незначна теломеразна активність виявляється в клітинах з високою швидкістю оновлення (лімфоцити, клітини епідермісу шкіри, епітелію тощо).