БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 14. МЕТАБОЛІЗМ ЛІПІДІВ
14.3.Розщепленення вищих жирних кислот
У хребетних тварин не менше 50 % енергії, яка вивільнюється в процесах окиснення макромолекул у клітинах печінки, нирок, легенів, серцевого й скелетного м'язів, постачається за рахунок окиснення вищих жирних кислот (ВЖК). У тварин, які голодують або впадають у зимову сплячку, а також у птахів під час перельоту окиснення цих кислот є єдиним джерелом енергії.
Основним механізмом окиснення жирних кислот, що відбувається в мітохондріях, є P'-окиснення, тобто окиснення за другим (β) атомом вуглецю. Спочатку в ділянці зовнішньої мембрани мітохондрій вони активуються ацил-КоА-синтетазою за участю АТФ і цитоплазматичного КоА-SH:
або ферментом КоА-ліазою (ацилтіокіназою) з використанням ГТФ:
![]()
Продукти ферментативної активації ВЖК - ацил-КоА проникають через внутрішню мітохондріальну мембрану за допомогою системи перенесення, яка складається з трьох мембранозв'язаних ферментів і низькомолекулярної сполуки - карнітину (Р-гідрокси-γ-триметиламонійбутират) (рис. 14.4).
Карнітин - широко розповсюджена у тканинах сполука, особливо її багато у м'язах. Проникнення активованих коротколанцюгових ЖК у мітохондрії можливе без участі карнітину, але транслокація довголанцюгових ВЖК неможлива без утворення ацилкарнітинового похідного.
Перенесення залишку жирної кислоти від ацил-КоА на карнітин каталізується карнітинацилтрансферазою, локалізованою на зовнішній мембрані мітохондрій:
![]()
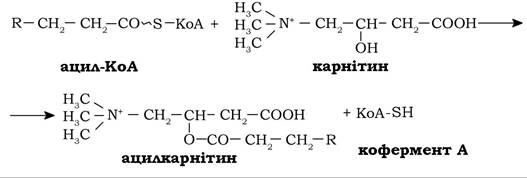
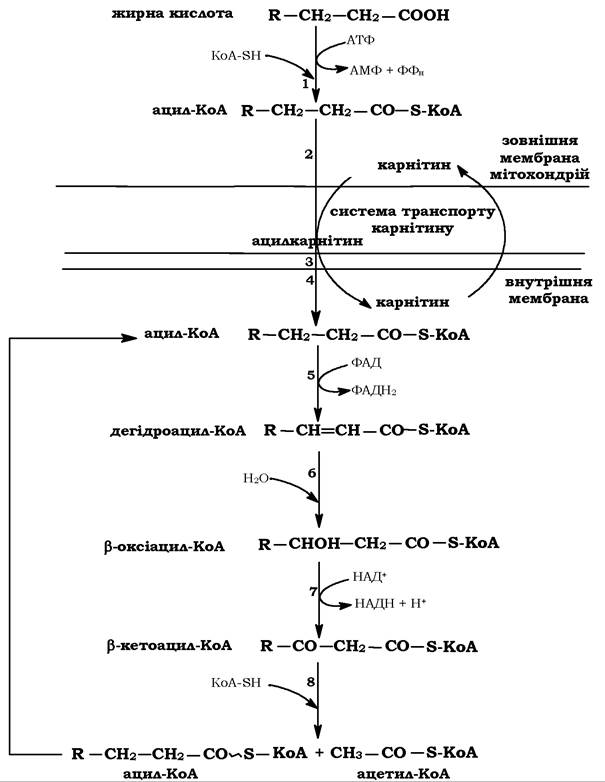
Рис. 14.4. Схема β-окиснення вільних жирних кислот:
1 - ацил-КоА-синтетаза; 2 - карнітинацилтрансфераза І; 3 - карнітин- ацилтранслоказа;
4 - карнітинацилтрансфераза ІІ; 5 - ацил-КоА-дегідрогеназа; 6 - β-оксіацил-КоА-гідратаза;
7 - β-оксіацил-КоА-дегідрогеназа; 8 - тіолаза (ацетил-КоА-ацилтрасфераза)
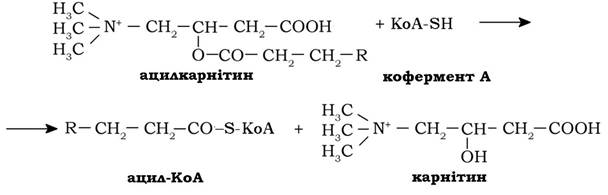
Далі ацилкарнітин переноситься транслоказою через внутрішню мітохондріальну мембрану.
У матриксі мітохондрій залишок жирної кислоти від ацилкарнітину за участю ацилтрансферази ІІ переноситься на внутрішньомітохондріальний КоА-SH (попередня реакція проходить у зворотному напрямку):
Після транслокації ацил-КоА в матрикс мітохондрій починається процес β-окиснення, що здійснюється циклічно чотирма послідовними ферментативними реакціями. При цьому кожен цикл реакцій закінчується вкороченням молекули ацил-КоА на два атоми вуглецю в результаті відщеплення молекули ацетил-КоА.
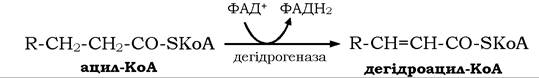
Спочатку ацил-КоА за участю специфічних ФАД-вмісних ферментів ацил-КоА-дегідрогеназ дегідрується з відщепленням двох атомів водню від карбоксильного кінця молекули ацил-КоА:
Далі дегідроацил-КоА за дії ферменту β-оксіацил-КоА-гідратази (енол-КоА-гідратази) перетворюється на β-оксіацил-КоА:
![]()
У наступній реакції β-оксіацил-КоА дегідрується НАД+-залежною 3-гідроацил-КоА-дегідрогеназою з утворенням β-кетоацил-КоА:
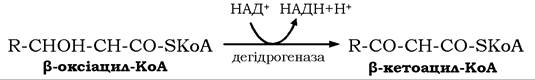
Остання стадія окиснення ВЖК каталізується β-кетоацилтіолазою (тіолазою). У цій реакції β-кетоацил-КоА взаємодіє з вільним КоА-SH і розщеплюється з утворенням молекул ацетил- КоА та ацил-КоА, укороченою на два атоми вуглецю:
![]()
Такі цикли реакцій β-окиснення повторюються доти, поки ацил-КоА повністю не перетвориться на вільний ацетил-КоА. Отже, окиснення жирної кислоти, яка містить 2п атомів вуглецю, здійснюється за (п - 1) циклів.
β-Окиснення ВЖК з непарною кількістю атомів вуглецю відбувається аналогічним чином, але в кінці процесу окиснення залишається одна молекула пропіоніл-КоА (CH3-CH2-CO~S-KoA), яка перетворюється в сукциніл-КоА, що окиснюється в циклі трикарбонових кислот.
Окиснення ненасичених ВЖК відбувається з певними особливостями, які полягають в тому, що утворені цис-ацил-КоА-похідні ізомеризуються під дією цис-транс-еноїл-КоА-ізомерази в дегідроа-цил-КоА, який вступає в подальшому у звичайну послідовність реакцій β-окиснення.
β-Окиснення ВЖК з довгим вуглецевим ланцюгом (С20, С22) можливе також у пероксисомах, де продуктами окиснення є ацетил-КоА і Н2О2. Воно включається при збагаченій жирами дієті, при вживанні гіполіпідемічних лікарських препаратів та ін.
Як уже відмічалося, процес β-окиснення ВЖК є основним шляхом катаболізму жирних кислот. Проте в ендоплазматичному ретикулумі може проходити процес ω-окиснення, де кінцева метильна група перетворюється в оксиметильну, а вона, у свою чергу, окиснюється до карбоксильної. Наслідком цього є утворення дикарбонової кислоти, яка в процесі β-окиснення розщеплюється до суберинової (С8) та адипінової (С6) кислот, які виводяться із сечею.
У клітинах мікроорганізмів, рослин і тварин виявлено процес α-окиснення ВЖК (відщеплення одновуглецевих фрагментів від карбоксильного кінця молекули жирної кислоти). У цьому випадку за дії мітохондріальної монооксидази ВЖК гідроксилюються по α-атому вуглецю з наступним окисненням і декарбоксилюванням в ендоплазматичному ритикулумі.
Енергетичне значення β-окиснення вільних жирних кислот. Наслідком проходження одного циклу β-окиснення ВЖК є утворення однієї молекули ацетил-КоА, при окисненні якої в циклі трикарбонових кислот утворюється три молекули НАДН і одна молекула ФАДН2. Крім того, у циклі β-окиснення ВЖК утворюється ще одна молекула ФАДН2 і одна молекула НАДН. Таким чином, при повному окисненні однієї молекули ВЖК, яка містить 2n атомів вуглецю, утворюється n молекул ацетил-КоА, які приводять до утворення в процесі окисного фосфорилювання 12n молекул АТФ і (n - 1) молекул ФАДН2 і НАДН, які разом сприяють утворенню 5(n - 1) молекул АТФ. Але 1 молекула АТФ використовується в процесі активації жирної кислоти. Отже, розщеплення однієї молекули жирної кислоти, яка містить 2n атомів вуглецю, приводить до утворення 12n + 5(n - 1) -1 = 17n - 6 молекул АТФ. Для порівняння відмітимо, що окиснення ВЖК, яка містить 6 атомів вуглецю (С6), приводить до утворення 45 молекул АТФ, тоді як повне окиснення глюкози, яка також має 6 атомів вуглецю, забезпечує енергією синтез тільки 38 молекул АТФ.
Регуляція процесу β-окиснення жирних кислот. Інтенсивність β-окиснення ВЖК залежить насамперед від активності регуляторного ферменту процесу - карнітинацилтрансферази І, яка контролюється алостеричним інгібітором ферменту - малоніл-КоА. Висока концентрація малоніл-КоА в цитозолі в абсорбтивний період стимулює біосинтез ВЖК і одночасно гальмує швидкість β-окиснення цих кислот. Однак у постабсорбтивний період, під час мобілізації жирних кислот із жирових депо й високої концентрації їх у крові, синтез малоніл-КоА припиняється. За цих умов підвищення активності карнітинацилтрансферази у випадку відсутності інгібітору стимулює β-окиснення вільних (мобілізованих) жирних кислот.