БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 15. ІНТЕГРАЦІЯ МЕТАБОЛІЧНИХ ШЛЯХІВ. ГОРМОНИ
15.1.Компартменталізація метаболічних шляхів
У клітинах контроль за етапами метаболізму здійснюється шляхом розділення метаболічних процесів по окремих відсіках (компартментах) (табл. 15.1):
✵ метаболічні перетворення й біосинтез в основному пов'язані з цитозолем. НАДФН, необхідний для реакцій відновлення, утворюється також у цитозолі в результаті окисного пентозофосфатного шляху глюкози;
✵ окисне розщеплення, пов'язане з диханням, відбувається в мітохондріях. Як коферменти зазвичай використовують НАД+ і флавопротеїни.
Таблиця 15.1
Компартменталізація деяких основних метаболічних шляхів
Компартмент |
Метаболічний процес |
Цитозоль |
Гліколіз Глюконеогенез Пентозофосфатний шлях Біосинтез ліпідів Біосинтез пуринів і піримідинів |
Мітохондрія |
Цитратний цикл β-окиснення жирних кислот Синтез кетонових тіл Дихальний ланцюг |
Інтеграція метаболічних шляхів забезпечує життєдіяльність організму:
✵ виробництво енергії у процесі окисного розпаду харчових речовин (механізми вивільнення й використання енергії були описані раніше);
✵ біосинтез речовин і структур. Виробництво ключових метаболітів і відновлених коферментів, необхідних для біосинтезу, забезпечується інтеграцією метаболічних шляхів.
У більшості випадків єдиним донором водню для різних біосинтезів є НАДФН, що теж є проявом інтеграції.
Ключові метаболіти, що містяться в точках розгалуження метаболічних шляхів, забезпечують перемикання метаболізму з одного шляху на інший залежно від потреб організму. Ці питання частково розглядалися раніше, тому наведемо лише декілька прикладів.
Глюкозо-6-фосфат утворюється в точці з'єднання й розгалуження таких процесів, як депонування глюкози - синтез глікогену, використання запасів енергетичного матеріалу - розпад глікогену, синтез НАДФН - пентозофосфатний шлях, вироблення енергії - аеробний розпад глюкози.
Піруват починає загальний для вуглеводів, білків і жирів шлях катаболізму і відкриває для них вхід у цитратний цикл, де утворюються метаболіти, які використовуються для глюконеогенезу й синтезу ряду амінокислот. Піруват також може бути субстратом для цих синтезів.
Ацетил-СоА включає білки, жири й вуглеводи в першу реакцію цитратного циклу. Він також служить субстратом для синтезу кетонових тіл. Крім того, у період травлення ацетил-СоА експортується в цитозоль у формі цитрату і використовується для синтезування жирних кислот. Особливість метаболізму жирних кислот полягає в необоротності перетворення пірувату на ацетил-СоА і, відповідно, неможливості перетворення жирних кислот на глюкозу.
Регуляторні механізми перемикання одного метаболічного шляху на інший частково розглядалися раніше. Отже, регуляція включає загальні моменти: зміну концентрації ферментів; зміну активності ферментів; зміну концентрації субстратів, продуктів, а також кофакторів. Як приклад, розглянемо регуляцію метаболізму вуглеводів, жирів і білків під впливом інсуліну та глюкагону. Перемикання метаболічних шляхів під дією цих гормонів відбувається постійно залежно від ритму харчування, складу їжі та фізіологічної активності організму.
У період травлення вплив гормонів направлений на накопичення речовин-енергоносіїв для використання їх у подальшому. Після надходження з кишечнику глюкози, її концентрація в крові збільшується, і це є стимулом для секреції інсуліну. Останній стимулює процеси депонування всіх мономерів, що утворилися в результаті травлення. Так, інсулін прискорює синтез глікогену в печінці та м'язах, синтез ліпідів і білків. Інсулін збільшує поглинання глюкози деякими клітинами й таким чином прискорює шляхи її використання (рис. 15.2).
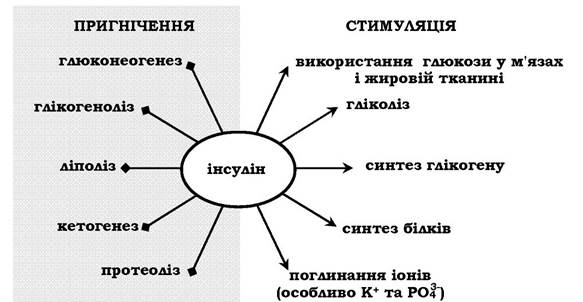
Рис. 15.2. Дія інсуліну
У постабсорбативний період концентрація глюкози знижується. У результаті знижується секреція інсуліну й зменшується поглинання глюкози всіма тканинами, крім нервової. Низька концентрація глюкози у крові є сигналом для секреції глюкагону й кортизолу.
Глюкагон прискорює розпад глікогену та мобілізацію триацил- гліцеринів, переключаючи окиснення тканинами переважно глюкози на окиснення жирних кислот і кетонових тіл. Глюкагон і кортизол прискорюють глюконеогенез із амінокислот і гліцерину для підтримання концентрації глюкози у крові на постійному рівні, потрібному для забезпечення мозку (рис. 15.3).
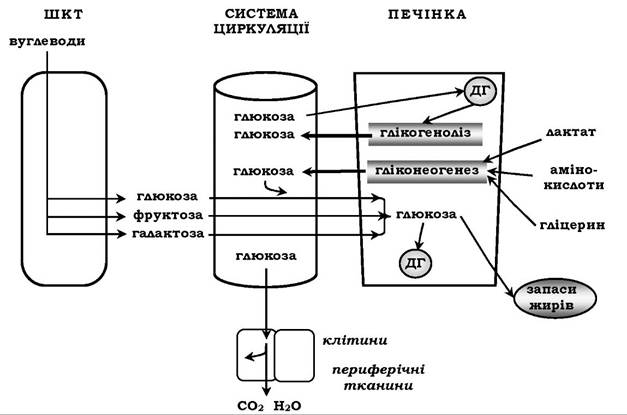
Рис. 15.3. Гомеостаз глюкози. ДГ - депо глікогену
У таблиці 15.2 показано вплив основних гормонів на обмін речовин. Далі наведено короткий опис деяких інших гормонів, що беруть участь в обміні вуглеводів, ліпідів і амінокислот
Обмін вуглеводів. Інсулін збільшує частку участі глюкози в процесах утворення енергії при незмінному загальному рівні енергопродукції. Активація інсуліном глікогенсинтетази і гліко-генрозгалужувального ферменту сприяє збільшенню синтезу глікогену. Поряд з цим інсулін виявляє інгібуючий вплив на глюкозо-6-фосфатазу печінки й гальмує таким чином вихід вільної глюкози в кров. Кінцевим результатом дії інсуліну (у разі його надлишку) є гіпоглікемія, що стимулює секрецію гормонів-
антагоністів інсуліну, до яких належать адреналін, норадреналін, глюкагон, соматотропін, глюкокортикоїдні й тиреоїдні гормони. У разі відносної чи абсолютної інсулінової недостатності порушуються процеси надходження глюкози до інсулінозалежних тканин, знижується окисне фосфорилювання та утворення глю- козо-6-фосфату; далі порушуються гліколітичне окиснення глюкози, цикл Кребса і гексозомонофосфатний (пентозний) цикл, пригнічується синтез глікогену й посилюється глікогеноліз.
Таблиця 15.2
Будова, механізми дії та основні ефекти гормонів, що регулюють метаболізм вуглеводів, білків і жирів
Будова |
Сигнал для секреції |
Органи- мішені |
Механізм передачі сигналу |
Зміна метаболізму у клітинах-мішенях |
Інсулін (місце синтезу - fi-клітини підшлункової залози) |
||||
Білок |
↑ Концентрація глюкози |
Печінка |
Через мембранні рецептори |
1) Прискорення синтезу глікогену; 2) прискорення синтезу білка; 3) гальмування глюконеогенезу. |
М'язи |
1) Прискорення синтезу глікогену; 2) прискорення синтезу білка; 3) прискорення транспорту глюкози у клітину. |
|||
Жирова тканина |
1) Прискорення синтезу жирів із глюкози; 2) прискорення транспорту глюкози у клітину |
|||
Глюкагон (α-клітини підшлункової залози) |
||||
Пептид |
↓ Концентрація глюкози |
Печінка |
Через мембранні рецептори |
1) Прискорення розпаду глікогену; 2) прискорення глюконеогенезу. |
Жирова тканина |
1) Прискорення ліполізу |
|||
Адреналін (клітини мозкового шару надниркових залоз) |
||||
Похідне тирозину |
Сигнал ЦНС |
Печінка |
Через мембранні рецептори |
1) Прискорення розпаду глікогену |
М'язи |
1) Прискорення розпаду глікогену |
|||
Жирова тканина |
1) Прискорення ліполізу |
|||
Кортизол (клітини коркового шару надниркових залоз) |
||||
Стероїд |
Концентрація глюкози в крові, опосередкована кортикотропіном |
Печінка |
Через плазматичні рецептори |
1) Прискорення глюконеогенезу; 2) індукція синтезу ферментів глюконеогенезу й катаболізму амінокислот. |
М'язи |
1) Прискорення катаболізму амінокислот; 2) зниження кількості надходження амінокислот. |
|||
Катехоламіни стимулюють глікогеноліз у печінці та м'язах. Збільшення синтезу сАМР під дією катехоламінів і більшою мірою адреналіну активує фосфорилазу печінки, розпад глікогену й утворення більшої кількості вільної глюкози. При цьому збільшується поглинання кисню, витрати енергії у зв'язку з посиленням серцевої діяльності, підвищенням м'язового тонусу й окисненням молочної кислоти в печінці. Глюкагон подібно до адреналіну активує аденілатциклазу, утворення сАМР, фосфорилазу, глікогеноліз і вихід глюкози з печінки у кров'яне русло. Цей вплив набагато сильніший, ніж у адреналіну, але глюкагон не діє на м'язову фосфорилазу, а отже, не мобілізує глікоген м'язів. Гіперглікеміч- ний ефект глюкагону є результатом стимуляції печінкового глікогенолізу та глюконеогенезу, індукції секреції адреналіну, гальмування проникнення глюкози у м'язи.
Гормон росту збільшує вихід глюкози в печінкові вени, посилює глюконеогенез, зменшує поглинання глюкози на периферії та посилює ліполіз, у результаті чого у крові підвищується концентрація вільних жирних кислот, які пригнічують дію інсуліну на мембранний транспорт глюкози.
Глюкокортикоїди стимулюють катаболізм білків і глюконеогенез, підвищують вміст глікогену в печінці й дещо меншою мірою у м'язах, зменшують мембранний транспорт глюкози та її утилізацію на периферії. Гіперглікемічна дія АКТГ опосередковується головним чином через глюкокортикоїди.
Обмін ліпідів. Мембрана адипоцитів містить рецептори, що взаємодіють з гормонами, яким притаманні ліполітичні властивості (катехоламіни, АКТГ, СТГ), та інсулінові рецептори. У результаті дії ліполітичних гормонів підвищується активність аденілатциклази, збільшується утворення сАМР, активізується ліпопротеїнлі- паза й ліполіз жиру. Взаємодія інсуліну з відповідними рецепторами, навпаки, призводить до гальмування ліполізу, який збільшується під час голодування, при тривалій роботі, охолодженні, стресі. Ліполітична дія катехоламінів (адреналін, норадреналін) і глюкагону здійснюється шляхом активації аденілатциклази. Роль норадреналіну в процесі ліполізу є важливішою, ніж адреналіну. Він утворюється в нервових адренергічних закінченнях у жировій тканині й забезпечує мобілізацію жирних кислот.
Гормон росту виявляє потужну ліполітичну дію, яка відрізняється від дії катехоламінів. Він викликає збільшення концентрації вільних жирних кислот у плазмі через 2-3 години. Ця дія, що спостерігається навіть при введені невеликих доз соматотропіну, напевне, пов'язана з гальмуванням процесу реетерифікації вільних жирних
кислот. Тим не менш соматотропін має певний модулюючий вплив і на активність аденілатциклази. Інші гіпофізарні гормони (АКТГ, ТТГ, меланоцитостимулюючий гормон) також виявляють певну ліпо- літичну дію, хоча й менш виражену, ніж соматотропін.
Інсулін має характерну антиліпотичну властивість, і при цукровому діабеті внаслідок збільшення ліполізу підвищується концентрація вільних жирних кислот у плазмі.
Обмін амінокислот. Анаболітична дія інсуліну полягає у прискорені проникнення амінокислот через мембрану клітини та включення їх у білки, що викликає зниження рівня амінокислот в крові. Інсулін знижує активність амінотрансфераз і ферментів циклу сечовини.