БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 3. КЛІТИНА І ПОЗАКЛІТИННИЙ МАТРИКС
3.1. Молекулярна й надмолекулярна організація клітини
3.1.2. Клітини еукаріотів
Організми надцарства еукаріотів складаються з еукаріотичних клітин, головною відмінністю яких від прокаріотичних є наявність ядра - мембранного компартмента, що містить більшу частину клітинної ДНК, і відділеного подвійною мембраною від цитоплазми (рис. 3.2). Ця структура має кулеподібну, яйцеподібну та інші форми діаметром від 0,5 мкм (у грибів) до 50 мкм (деякі яйцеклітини). Головні функції клітинного ядра: 1) зберігання інформації у формі ДНК; 2) передача інформації в цитоплазму шляхом транскрипції - синтез РНК, які забезпечують перенесення інформації; 3) передача інформації дочірнім клітинам при реплікації - поділ клітин і ядер. Ядро складається з: 1) нуклеоплазми - рідкої частини, ядерного матриксу (тривимірний каркас із білків) і різних включень; 2) хромосом, кожна з яких містить дві хром.атиди - нуклеопротеїдні структури з однієї молекули ДНК і білків; 3) ядерної оболонки з двох мембран (з порами до 100 нм), між якими знаходиться перинуклеарний простір (10-40 нм). В ядрах є ущільнені ділянки - ядерця, де здійснюється синтез рРНК (рибосомальної РНК) на ядерцевій ДНК.
У цитоплазмі локалізовано багато органоїдів, які виконують певні специфічні функції. Одна з таких структур - мітохондрія - досить цікава тим, що дуже схожа на прокаріотичні клітини: за формою, за розміром, містить ДНК і рибосоми, синтезує АТФ (аденозинтрифосфат) - універсальне джерело енергії в клітині, розмножується бінарним поділом. До інших функцій мітохондрій відносять біосинтетичні процеси (синтез білків, стероїдних гормонів), транспорт іонів. Як і ядро, мітохондрії складаються із двох м,ембран з міжмембранним простором (перимітохондріальним) і матриксу, що містить продукти обміну, ферменти циклу лимонної кислоти, окиснення жирних кислот тощо.
Як "нащадків" прокаріотичних клітин можна розглядати й хлоропласти, котрі здійснюють фотосинтез органічних речовин за рахунок енергії сонячного світла, що поглинається хлорофілом. За розмірами, організацією хлорофіловмісних мембран, розмноженням поділом, нуклеотидною послідовністю ДНК - хлоропласти дуже схожі з ціанобактеріями (синьозелені водорості). Це також указує на те, що хлоропласти, як і мітохондрії, походять від прокаріотів.
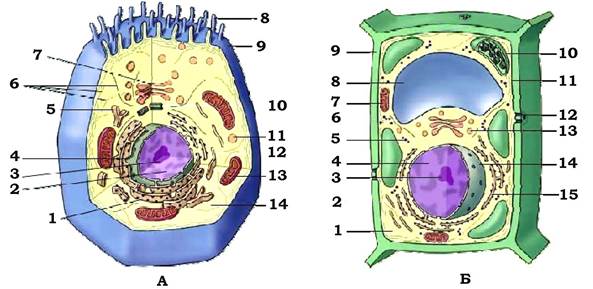
Рис. 3.2 Схема еукаріотичної клітини з основними органоїдами:
А - тваринна клітина: 1 - ендоплазматичний ретикулум (гранулярний);
2 - ядро; 3 - ядерце; 4 - ядерна оболонка; 5 - ендоплазматичний ретикулум (гладенький); 6 - цитоскелет; 7 - апарат Гольджі; 8 - мікроворсинки; 9 - плазматична мембрана; 10 - центріолі; 11 - лізосоми; 12 - рибосоми; 13 - мітохондрії; 14 - цитоплазма.
Б - рослинна клітина: 1 - цитоплазма; 2 - ендоплазматичний ретикулум (гладенький); 3 - ядерце; 4 - ядро; 5 - вільні рибосоми; 6 - апарат Гольджі; 7 - мітохондрії; 8 - центральна вакуоль; 9 - стінка клітини; 10 - хлоропласти; 11 - плазматична мембрана; 12 - плазмодесма; 13 - лізосоми; 14 - оболонка ядра; 15 - ендоплазматичний ретикулум (гранулярний)
Еукаріотична клітина від прокаріотичної (табл. 3.1) відрізняється ще й тим, що містить багато внутрішньоклітинних мембран, які розділяють клітину на окремі компартменти. Далі буде показано, що в еукаріотичній клітині в більшості (якщо не в усіх) процесів прямо чи опосередковано беруть участь мембрани. Це потребує і значного збільшення поверхні внутрішньоклітинних мембран за рахунок складок, вигинів, везикул та інших їхніх форм. Найбільше в клітині мембран ендоплазматичного ретикулума (ЕР), на якому синтезуються ліпіди й мембранні білки, "створюється" матеріал для "експорту" з клітини. Такий експорт - транспорт речовин із клітини здійснюється в результаті екзоцитозу: внутрішньоклітинні мембранні везикули зливаються з плазматичною мембраною й вивільняють у зовнішнє середовище свій вміст. Зворотним процесом - ендоцитозом - у клітину потрапляють речовини з позаклітинного матриксу. У деякій літературі згадуються мікросоми. Слід пам'ятати, що мікросом в інтактних клітинах немає. Ці везикули спонтанно утворюються переважно з ендоплазматичного ретикулума в результаті гомогенізації клітин і тканин. ЕР складається зі сплющених цистерн, покритих рибосомами (шорсткий, гранулярний ЕР) чи без них (гладенький, агранулярний ЕР).
Рибосома складається з двох рибонуклеопротеїдних субоди- ниць, вони можуть вільно локалізуватися в цитозолі чи приєднуватися до ЕР через білки, які вони (рибосоми) синтезують. У процесі синтезу білка вздовж мРНК може переміщуватися багато рибосом - такі ланцюги рибосом називаються полірибосо- мами (полісомами). Внутрішньоклітинні мембрани утворюють і апарат Гольджі, відкритий у 1898 р. - це стопки сплющених мембранних пухирців (диктіосоми), які також сприяють синтезу і транспорту органічних молекул - білків, вуглеводів, ліпідів. Ця функція особливо виражена в секреторних клітинах. Апарат Гольджі виконує ще одну функцію - формування лізосом.
Лізосоми також являють собою мембранні везикули (0,20,5 мкм), що містять необхідні для внутрішньоклітинного травлення ферменти. Лізосомні ферменти синтезуються рибосомами на гранулярному ЕР, транспортуються до апарату Гольджі, від якого пізніше відбруньковуються мембранні пухирці з "дозрілими" ферментами й утворюють ендолізосоми (рис. 3.3).
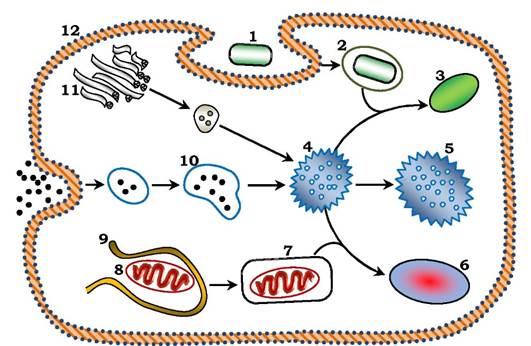
Рис. 3.3. Три шляхи утворення лізосом (схема):
1 - бактерія; 2 - фагосома; 3 - фаголізосома; 4 - ендолізосома; 5 - лізосома; 6 - аутофаголізосома; 7 - аутофагосома; 8 - мітохондрія; 9 - ендоплазматичний ретикулум; 10 - ендосома; 11 - апарат Гольджі; 12 - плазматична мембрана
Лізосоми утворюються шляхом злиття ендолізосом з ендоцитозними пухирцями (ендосомами). Друга різновидність лізосом - аутофаголізосоми, які є результатом злиття аутофагосоми з ендолізосомою. При злитті фагосоми з ендолізосомою утворюється фаголізосома. Продукти перетравлення трьох типів лізосом засвоюються цитоплазмою клітини, а лізосома, що містить непере- травлений матеріал, називається залишковим тільцем. Ці структури можуть залишатися в клітинах (гепатоцитах та ін.) або їхній вміст виводиться шляхом екзоцитозу. Ще донедавна традиційно виявляли первинні та вторинні лізосоми. (На жаль, такий поділ наведено і в деяких сучасних підручниках). Експериментальні дослідження останніх десятиріч свідчать про те, що лізосомні гідролази та мембранні білки лізосом відбираються за допомогою різних рецепторів. Вони залишають апарат Гольджі в окремих транспортних пухирцях і вперше зустрічаються в ендолізосомі, яка вже має субстрат для розщеплення.
Аналогічним способом утворюються пероксисоми (мікротільця), в яких розкладаються токсичні пероксиди, що утворились у процесі клітинного дихання:
![]()
Ці мікротільця мають сферичнуформу з діаметром 0,3-1,5 мкм, походять від ЕР, з яким зв'язок іноді не втрачається. Пероксисоми рослинних клітин поділяють на гліоксисоми (беруть участь у перетравленні ліпідів), пероксисоми листків (нейтралізують пероксид водню - один із продуктів фотодихання) і неспеціалізовані пероксисоми, котрі виявляються в інших тканинах. Мембрани утворюють у рослині великі, заповнені рідиною везикули - вакуолі, а також інші постійні та тимчасові мембранні утворення.
Усі ці мембранні структури займають майже половину об'єму клітини й відповідають певним компартментам. Частина цитоплазми, позбавлена всіх органоїдів (органел), називається цитозолем.
Особливе місце серед клітинних мембран посідає плазматична мембрана (ПМ). Загалом вона нагадує плазматичну мембрану прокаріотичної клітини, проте за молекулярною організацією і функціями значно відрізняється. Цілком можливо, що висока спеціалізація плазматичної мембрани еукаріотичної клітини почалася з того моменту, коли функція утворення АТФ перейшла від ПМ у прокаріотів до мітохондрій в еукаріотів. У плазматичній мембрані локалізуються іонні насоси, завдяки яким клітина може в межах мілісекунд змінити внутрішньоклітинну концентрацію основних неорганічних іонів (K+, Na+, Ca2+) і свою проникність, що веде до зміни потенціалу на мембрані. Це є визначальним для передачі електричних сигналів (наприклад, по нервовій системі). Завдяки унікальній молекулярній організації плазматична мембрана еукаріотів першою сприймає, класифікує та трансформує енергію зовнішніх стимулів (світло, температура, гормони, отрути, ліки) в енергію біологічного збудження.
Функцію контролю положення і руху описаних органоїдів здійснює цитоскелет, який складається з мікротрубочок, мікрофіламентів і проміжних філаментів. Мікротрубочки - тонкі трубочки діаметром 24 нм, товщина стінок яких становить 5 нм, довжина - декілька мікрометрів, сформовані з глобулярного білка тубуліну. Мікротрубочки виконують структурну роль - утворюють опорну систему клітини (звідси назва - цитоскелет), підтримують форму клітин (наприклад, за дії колхіцину, який руйнує мікротрубочки, тваринні клітини набувають сферичної форми), сприяють переміщенню пухирців Гольджі, лізосом, мітохондрій. Із мікротрубочок побудовані центріолі (дрібні порожнисті циліндри довжиною 0,3-0,5 мкм і діаметром 0,2 мкм). При поділі клітин центріолі подвоюються і дві нові пари розходяться до полюсів веретена поділу (структура, по екватору якої локалізуються готові до розходження хромосоми), що також складається з мікротрубочок. За будовою до центріолей подібні базальні тільця, які розташовуються біля основи джгутиків та війок і побудовані з мікротрубочок. Рух веретена, війок і джгутиків здійснюється за рахунок ковзання мікротрубочок, які забезпечують розходження хромосом, биття джгутиків і війок.
Мікрофіламенти - тонкі нитки діаметром 5-7 нм, формуються з білка актину. Мікрофіламенти, як і мікротрубочки, утворюють сплетення, які нерідко прилягають до плазматичної мембрани, беруть участь в ендо- й екзоцитозі, а також у русі інших органоїдів. Мікрофіламенти складаються з актину й міозину. Показовим у цьому плані є функціонування мікроворсинок - пальцеподібних виростів плазматичної мембрани клітин усмоктувального типу (епітелій тонкого кишечнику та звивисті канальці нефронів). В основі кожної мікроворсинки пучки актинових ниток зв'язані з міозиновими. У результаті ковзних рухів актинових ниток уздовж міозинових мікроворсинка може поперемінно вкорочуватись і видовжуватись, що, імовірно, сприяє всмоктуванню речовин у клітину. Мікрофіламенти й асоційовані з ними білки під ПМ утворюють клітинний кортекс, який забезпечує міцність поверхні клітини, можливість зміни форми клітини й руху. Властивості клітинного кортексу визначаються балансом кооперативних і конкурентних взаємодій актинозв'язувальних білків (міозин, тропоміозин, спектрин тощо). ПМ настільки міцно зв'язана з актиновим клітинним кортексом, що вважається єдиним функціональним (не структурним) утворенням. Філаменти беруть участь у зв'язуванні клітин через адгезивні з'єднання.
В утворенні цитоскелета беруть участь і проміжні філаменти, діаметром 8-10 нм (за товщиною вони займають проміжне місце між актиновими мікрофіламентами й мікротрубочками). Ці утворення, як правило, формують "кошик" навколо ядра, звідки досягають не тільки периферії клітини, але й можуть брати участь у з'єднанні клітин одна з одною за допомогою десмосом (структури міжклітинного контакту). Проміжні філаменти утворюються з фібрилярних білків, серед яких переважають кератини, десміни і віментин.
Крім органоїдів у клітинах є включення. До них належать ліпідні гранули чи крапельки, що містять різні ліпіди. У кінцевому результаті гранули оточуються поодиноким шаром ліпідів. При посиленому відкладанні нейтральних жирів ліпідні гранули збільшуються і тоді вони вже називаються жировими гранулами. Останні в жирових клітинах тварин зливаються в одну велику центральну краплю. Місцем накопичення вуглеводів у клітинах тварин є гранули глікогену. Аналогічні функції в клітинах рослин виконують зерна крохмалю, оточені мембраною. Готовий до виділення секрет у клітинах міститься в секреторних гранулах.
Секреторні включення (секрет) є продуктом синтетичної активності спеціалізованих секреторних клітин і зазвичай виділяються постійно або у відповідь на зовнішній стимул, що діє на клітину. Екскреторні включення не містять будь-яких ферментів або інших активних речовин. Це продукти метаболізму, що підлягають видаленню (екскреції) з клітин. Пігментні включення можуть бути екзогенні (каротин, барвники тощо) і ендогенні (гемоглобін, гемосидерин, меланін, ліпофусцин). їхня присутність у цитоплазмі може змінювати колір тканини, органу (тимчасово або постійно). Часто пігментація тканини є діагностичною ознакою деяких патологій.
Отже, основною структурно-функціональною одиницею живих організмів є клітина (рис. 3.2). Вона є елементарною живою системою і може існувати як окремий організм, так і в складі багатоклітинного організму. Вмістом клітини є протоплазма (цитоплазма + ядро). Генетичний апарат у клітинах еукаріотів локалізований у ядрі, а в клітинах прокаріотів - у нуклеоїді (відповідає одній молекулі ДНК, закріпленій в одній точці на внутрішньому боці плазматичної мембрани). Клітини здатні до самовідтворення різними способами.
За розмірами клітини суттєво відрізняються: від 0,1-0,25 мкм (у деяких бактерій) до 155 мм (яйце страуса в шкарлупі). Але діаметр більшості клітин варіює від 10 до 100 мкм. Виконання численних функцій клітинами забезпечується органоїдами, до яких належать ядро, хромосоми, рибосоми, мітохондрії, ЕР, апарат Гольджі, лізосоми, плазматична мембрана тощо. Форма живої клітини підтримується цитоскелетом, сформованим із мікро- трубочок, проміжних філаментів і мікрофіламентів. Характерною особливістю клітин є просторово-часова організація (компартмен- талізація) і принцип компактності (наприклад, 10-12 г ДНК яйцеклітини організму людини містить інформацію про всі його білки). Завдяки плазматичній мембрані клітина підтримує постійність внутрішньоклітинного середовища, здатність до екзо- та ендоцитозу: може захоплювати не лише крапельки з великими молекулами (включаючи й білки), а й віруси. Окремі клітини (макрофаги, нейтрофіли) поглинають і бактерії.
Клітини еукаріотів мають схожий набір органоїдів, схожі механізми регуляції метаболізму, синтезу й гідролізу важливих макромолекул, запасання і використання енергії. У клітин про- та еукаріотів аналогічно використовується генетичний матеріал для синтезу білків, функціонує плазматична мембрана, багато спільних ознак свідчить про єдність їхнього походження. Разом із тим, різні клітини одного організму різняться формою, розмірами, кількістю органоїдів, набором ферментів тощо - це визначено виконанням специфічних функцій. Відмінності клітин багатоклітинного організму зумовлені різною активністю генів, що визначає різну диференціацію клітин. У результаті одні клітини проводять збудження (нервові), інші набувають властивостей скорочення (м'язові), треті синтезують гормони і травні ферменти (залозисті), четверті вкривають тіло й вистеляють порожнини організму (епітеліальні) тощо. Окрім того, багато клітин виконують не одну функцію, їх відносять до полі функціональних. Такими є, наприклад, гепатоцити. Вони синтезують білки, жовчні кислоти, накопичують глікоген, перетворюють глікоген на глюкозу, нейтралізують отрути та ін. Однак схожих ознак між клітинами більше, ніж спеціалізованих. Клітини мають і різний час життя. Скажімо, організм людини складається з 1014 клітин. Щоденно з цієї кількості гине й знов утворюється 7 · 1010 клітин кишкового епітелію і 2 · 109 еритроцитів. Тобто час життя клітин багатоклітинного організму коливається від 1-2 днів (епітелій кишечника) до кінця життя організму (нейрони, волокна скелетних м'язів).
Що ж стосується одноклітинних організмів, то відмінності у структурі пояснюються їхнім пристосуваннями до середовища існування. Більш того, у різних одноклітинних могли бути різні прокаріотичні попередники. Про розвиток клітин існує багато гіпотез, але найбільш реальними є дві. Гіпотеза симбіогенезу передбачає, що одні прокаріоти всередині клітини-господаря перетворилися в мітохондрії, інші - у хлоропласти, не втратили здатності до самовідтворення, але не як клітини, а як органоїди. Інша гіпотеза дотримується поступового розвитку власних структур прокаріотичної клітини в процесі перетворення її в клітину еукаріотичну. Усі клітини мають універсальну систему регуляції та саморегуляції: метаболіти й неорганічні іони всередині клітини діють або на ген, або на сам фермент. Унаслідок такої регуляції підтримується оптимальний динамічний рівень внутрішньоклітинних процесів. Велика кількість клітин, об'єднаних регуляторними процесами метаболізму й міжклітинними контактами, забезпечують надійність функціонування тканин і органів багатоклітинного організму.
Щодо організації клітин у тканини слід відмітити важливу роль позаклітинного матриксу, що включає різноманітні полісахариди й білки, організовані в сіткоподібні структури. Такий матрикс може утворювати структури кістки чи зуба, формувати прозору речовину рогівки, базальну мембрану та ін. Він впливає на розвиток клітин, їхню міграцію, проліферацію, форму, метаболізм. Останнє забезпечується тим, що водна фаза полісахаридного гелю забезпечує дифузію поживних речовин, метаболітів і гормонів між кров'ю та клітинами тканини.
Отже, клітини вперше відкрив Р. Гук у 1665 р. і лише півтора століття потому М. Шлейден і Т. Шванн сформулювали клітинну теорію, згідно з якою всі організми мають клітинну будову (18381839). Ще через 20 років (1858) Р. Вірхов обґрунтував принцип спадкоємності клітин шляхом поділу: "кожна клітина із клітини». Завдяки наступним відкриттям основних органоїдів, мітотичного поділу, відкриттів молекулярної біології клітини сформувалося нинішнє уявлення про клітинний рівень організації живого. Сучасна клітинна теорія розглядає організм як складно організовану інтегровану систему, яка складається із взаємодіючих клітин Для організму характерні специфічні властивості, які не
є сумою властивостей її складових клітин. Але не тільки схожість будови клітин про- та еукаріотів підтверджує клітинна теорія. Це підтверджується як хімічним складом клітин і організмів, так і їхніми аналогічними метаболічними процесами. Клітини всіх живих систем складаються з близьких за будовою та властивостями білків, вуглеводів, нуклеїнових кислот, неорганічних іонів і води. Існування вірусів - своєрідних клітинних паразитів - також є доказом універсальності клітинної будови живого.
Отже, клітина - це структурна і функціональна елементарна одиниця живих організмів. Усі клітини обмежені плазматичною мембраною, мають цитоплазму та ядро (ядерну зону в прокаріотів). Розміри клітини (табл 3.1) щонайменше визначаються розмірами молекул, з яких вони побудовані, швидкістю трансмембранного переміщення поживних речовин і кисню та співвідношенням між площею поверхні й об'ємом клітини.
Для забезпечення життєдіяльності клітині необхідна мінімальна кількість біомолекул, тому меншими вони бути не можуть. Вони не можуть бути й великими, бо швидкість метаболічних процесів залежить від дифузії речовин у цитозолі. У великих клітинах це обмежило б можливості регуляції метаболізму. З іншого боку, оскільки площа плазматичної мембрани відносно велика порівняно з її об'ємом, у клітину проникає достатнє число молекул речовин. Зі збільшенням діаметра клітини відношення S/ V різко знижується (S = 4πr2 ; V = 4/3πr3).
Існують два типи клітин: про- і еукаріотичні. Перші - малі, не мають мембранних компартментів, їхній генетичний матеріал не обмежений мембраною. Прокаріотичні клітини швидко ростуть і діляться. Адекватним прикладом таких клітин є бактерія Е. соli - типова складова кишкової мікрофлори людини і тварин. Має форму паличок довжиною ~ 2 мкм, діаметром - 0,8 мкм і об'ємом 1 мкм3, з густиною ~ 1,1 г/см3 і масою ~ 700 · 109 Да (1 · 10-9 мг). Вона краще ніж інші бактерії вивчена на молекулярному рівні й стала дуже корисною для біохімічних досліджень.
Еукаріотичні клітини значно більших розмірів (табл. 3.1), їхній об'єм у 1 000 - 10 000 разів перевищує об'єм прокаріотичних клітин. Припускається, що мітохондрії та хлоропласти походять від прокаріотів. Важливою ознакою еукаріотичної клітини є лізосоми, в яких локалізовані деструктуруючі ферменти, і пероксисоми.
Дослідження сучасних організмів і всіх біомолекул дозволяє припускати, що розвиток автокаталітичних механізмів у їхніх клітинах розпочався з еволюції групи молекул - предків РНК, які могли каталізувати і власну реплікацію, і синтез поліпептидів. Після накопичення додаткових каталітичних білків, що сприяло розвитку складніших клітин, ДНК замінила РНК у ролі носія генетичної інформації.
Таблиця 3.1
Приблизні розміри деяких атомів, молекул і структур клітини
№ |
Назва |
Довжина (більший радіус), нм |
1 |
Водень |
0,037 |
2 |
Кисень |
0,066 |
3 |
Азот |
0,070 |
4 |
Вуглець |
0,077 |
5 |
Сірка |
0,104 |
6 |
Фосфор |
0,110 |
7 |
Вода |
0,140 |
8 |
Аланін |
0,5 |
9 |
Глюкоза |
0,7 |
10 |
Фосфатидилхолін |
3,5 |
11 |
Гемоглобін |
6,8 |
12 |
Рибосома E. coli |
18 |
13 |
Бактеріофаг E. coli ФХ 174 |
18 |
14 |
Рибосома еукаріотів |
30 |
15 |
Вірус поліомієліту |
30 |
16 |
Ядерні пори |
90 |
17 |
Міозин |
100 |
18 |
Бактеріофаг E. coli Т 4 |
200 |
19 |
Вірус віспи |
250 |
20 |
Вірус тютюнової мозаїки |
300 |
21 |
Мікоплазма (діаметр) |
330 |
22 |
Рикетсії |
750 |
23 |
Стафілокок |
1000 |
24 |
Мітохондрії гепатоцитів |
1500 |
25 |
E. coli |
2250 |
26 |
Еритроцит |
8000 |
27 |
Спірохета |
8300 |
28 |
Гепатоцит |
20000 |
29 |
Кореневий волосок |
1 000000 |
Важливо також, що відбір РНК за кількістю білків, які вони кодують, не міг розпочатись раніше, ніж з'явився замкнений об'єм - компартмент, тобто формування "предка" плазматичної мембрани було вирішальним для виникнення першої клітини. Основна роль у біохімічній еволюції клітинних мембран належить класу амфіпатичних молекул, які здатні спонтанно агрегувати. Як тільки молекула РНК опинилась у замкненому мембраною просторі, нуклеотидні послідовності РНК могли впливати на ознаки цілої клітини.