МИКРОБИОЛОГИЯ Учебное пособие - 2012
ГЛАВА 6. ВИРУСЫ
6.4. РАЗМНОЖЕНИЕ ВИРУЛЕНТНОГО И УМЕРЕННОГО БАКТЕРИОФАГОВ
Бактериофаги делят на зрелые, существующие вне клетки хозяина (они инертны и неподвижны); вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные, которые проникают в клетку и остаются в ней в латентной форме.
Цикл размножения вирулентного бактериофага в клетке хозяина, завершающийся ее лизисом, называется литическим циклом инфекции. Этот цикл состоит из следующих этапов: адсорбция, внедрение, внутриклеточное размножение, созревание и освобождение (рис. 22).
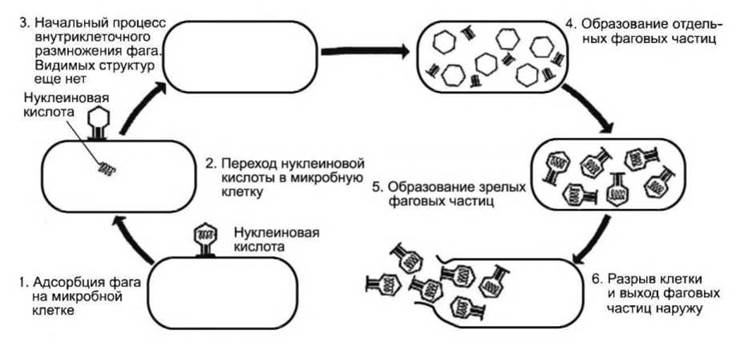
Рис. 22. Цикл развития вирулентного бактериофага
Адсорбция. Литический цикл инфекции начинается с того, что частица бактериофага случайно сталкивается с клеткой хозяина. Бактериофаги строго специфичны в отношении клетки хозяина. Специфичность отношений бактериальной клетки и фага определяется специфичностью адсорбции. Если у бактериофага участок адсорбции химически комплементарен специфическому рецепторному участку на поверхности бактериальной клетки, то происходит необратимая адсорбция.
У грамотрицательных бактерий рецепторы находятся в липопротеидном или в липополисахаридном слое клеточной стенки. У некоторых бактерий рецепторные участки локализованы на жгутиках или пилях.
У вирионов разных бактериофагов адсорбционные участки также различаются. У Т-четных фагов кишечной палочки органами адсорбции являются хвостовые нити, у других такими органами служит сам отросток. Во время адсорбции фаговая частица прикрепляется нитями к рецепторному участку клеточной стенки бактерии и фиксируется на ней при помощи шипов базальной пластинки.
Внедрение. После адсорбции фаговая частица инъецирует содержащуюся в ней ДНК внутрь бактериальной клетки. Содержащийся в отростке фаговый лизоцим разрыхляет клеточную стенку бактерии. Чехол отростка сокращается под действием фермента АТФ-азы, и полый стержень отростка проходит через клеточную стенку. Когда кончик стержня достигает клеточной мембраны, содержащаяся в головке бактериофага ДНК впрыскивается под клеточную стенку бактерии. Белковая оболочка капсида бактериофага, называемая «тенью», остается снаружи.
Такой механизм внедрения, характерный для Т-четных фагов кишечной палочки, является уникальным. Однако у некоторых фагов этот механизм совершенно другой. Так, у нитевидных фагов, содержащих однонитчатую ДНК, основной белок спирального капсида проходит через клеточную стенку бактерии и остается в клеточной мембране, а минорный белок оболочки проникает вместе с фаговой ДНК в цитоплазму.
Внутриклеточное развитие. Сразу после внедрения фаговая ДНК вызывает полную перестройку метаболизма зараженной клетки. Прекращается синтез бактериальной ДНК, подавляется синтез РНК и белков клетки. Затем синтез ДНК возобновляется с повышенной скоростью, причем сначала фаговая ДНК образуется за счет распавшейся бактериальной. Необходимые для синтеза фаговой ДНК ферменты, так называемые «ранние» белки, синтезируются сразу после заражения клетки. Затем начинается синтез «поздних» белков — субъединиц капсида и вирус-специфического белка — лизоцима. Одновременно с этим происходит конденсация молекул вирусной ДНК.
Во время перестройки обмена веществ инфицированной клетки никаких видимых изменений внутри нее не происходит, в связи с чем этот период называют скрытым или латентным.
Созревание и высвобождение. Каждая молекула ДНК плотно упаковывается в виде многогранника, затем с ней связываются субъединицы капсида и образуются головки фага. Процесс завершается сборкой головок и отростков в зрелые вирионы.
Фаговый лизоцим лизирует клеточную стенку бактерии, гидролизуя гликозидные связи в ее пептидогликановом слое. В результате клеточная стенка постепенно становится все менее прочной и, в конце концов, разрывается под действием внутриклеточного давления. Зрелые фаговые частицы выходят в окружающую среду и адсорбируются на молодых, размножающихся клетках бактерий. На этом литический цикл инфекции завершается.
Обычно фаги высвобождаются из клетки хозяина после ее лизиса. Однако нитевидные фаги являются исключением из этого правила. Созревание фага и его высвобождение происходят за счет того, что фаговая ДНК выталкивается из клетки и в цитоплазматической мембране соединяется с белком оболочки. В процессе высвобождения нитевидных фагов из клетки последняя не лизируется и остается жизнеспособной.
В отличие от вирулентного фага умеренные фаги не размножаются внутри бактериальной клетки и не лизируют ее. В зрелых фаговых частицах ДНК присутствует в виде линейной двойной спирали. После внедрения умеренного фага лямбда в клетку Е. coli его ДНК, называемая профагом, замыкается в кольцо. Кольцевая ДНК фага встраивается в хромосому бактерии и в течение длительного времени может реплицироваться вместе с ней и передаваться потомству. Клетки бактерий, несущие профаг, называются лизогенными. Лизогенная культура может содержать 2, 3 и более фагов. Она, как правило, устойчива к заражению теми фагами, которые присутствуют в ней в виде профага. Возникающий иммунитет обусловлен образованием особого белка-репрессора, подавляющего размножение вирулентного фага.
Лизогенная бактерия делится неограниченно долго, не подвергаясь лизису. Исключение профага из хромосомы бактерии и переход его в вегетативное состояние может произойти спонтанно или под воздействием мутагенных факторов (ультрафиолетовое или рентгеновское облучение, пероксид водорода и некоторые другие вещества). После исключения профаг снова становится автономным; с этого момента он перестает подчиняться репрессии и начинает размножаться в клетке как вирулентный фаг, что в конечном итоге приводит к лизису клетки хозяина и высвобождению фага.