БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 5. БУДОВА, ВЛАСТИВОСТІ Й ФУНКЦІЇ БІЛКІВ
5.5. Особливості функціонування олігомерних білків
5.5.2. Структура й функції гемоглобіну
Гемоглобіни - споріднені білки, які містяться в еритроцитах людини і хребетних тварин. Ці білки виконують дві важливі функції:
✵ перенесення О2 із легень до периферичних тканин;
✵ участь у перенесенні СО2 і протонів із периферичних тканин у легені для подальшого видалення з організму.
Кров щоденно повинна переносити з легень у тканини близько 600 л О2. Оскільки О2 погано розчинний у воді, то практично весь кисень у крові зв'язаний з гемоглобіном еритроцитів.
Від здатності гемоглобіну насичуватись О2 в легенях і відносно легко віддавати його в капілярах тканин залежить кількість отриманого тканинами О2 та інтенсивність метаболізму. З іншого боку, О2 - сильний окисник, надлишок надходження О2 у тканини може привести до пошкодження молекул і порушення структури та функцій клітин. Тому найважливішою функцією гемоглобіну є його здатність регулювати спорідненість до О2 залежно від умов у тканинах.
Гемоглобіни, як і міоглобін, відносять до гемопротеїнів, але вони мають четвертинну структуру (складаються з чотирьох поліпептидних ланцюгів), завдяки якій виникає можливість регуляції їхніх функцій. В еритроцитах дорослої людини гемоглобін становить 90 % від усіх білків даної клітини.
Гемоглобін А - основний гемоглобін дорослого організму, становить близько 98 % від загальної кількості гемоглобіну, тетрамер, складається з двох поліпептидних ланцюгів а і β (2α2β).
Гемоглобін А2 - знаходиться в організмі дорослої людини в меншій кількості, на його частку припадає близько 2 % від загального гемоглобіну. Він складається з 2α- та 2δ-ланцюгів.
Гемоглобін А1с - гемоглобін А, модифікований ковалентним приєднанням до нього глюкози (так званий глюкозований гемоглобін).
Ембріональний гемоглобін синтезується в ембріональному жовчному мішку через декілька тижнів після запліднення. Являє собою тетрамер 2ξ2ε. Через два тижні після формування печінки плоду в ній починає синтезуватися гемоглобін F, який до шести місяців заміняє ембріональний гемоглобін.
Гемоглобін F - фетальний гемоглобін, синтезується в печінці та в кістковому мозку плоду до періоду його народження. Має тетрамерну структуру, яка складається з двох α- та двох γ-ланцюгів. Після народження дитини поступово замінюється на гемоглобін А, який починає синтезуватись у клітинах кісткового мозку вже на восьмому місяці розвитку плоду.
Конформація окремих протомерів гемоглобіну нагадує конформацію міоглобіну, незважаючи на те, що в первинній структурі їхніх поліпептидних ланцюгів ідентичними є 24 амінокислотні залишки. Протомери гемоглобіну, як і апоміоглобін, складаються з восьми спіралей, скручених у щільну глобулярну структуру, яка містить внутрішнє гідрофобне ядро та "кишеню" для зв'язування гему. З'єднання гему з глобіном (білковою частиною) аналогічне такому ж у міоглобіні - гідрофобне оточення гему, за винятком двох залишків Гіс Е7 і Гіс F8 (рис. 5.31). Проте тетрамерна структура гемоглобіну порівняно з міоглобіном має складніший структурно-функціональний комплекс, ніж міоглобін.
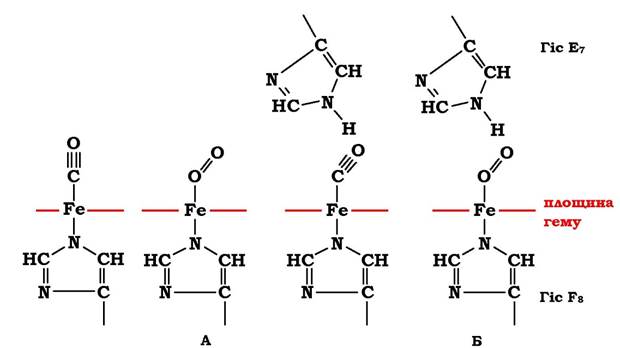
Рис. 5.31. Просторове розташування СО та О2, зв'язаних з вільним гемом (А) і гемом у складі гемоглобіну або міоглобіну (Б)
Гем має високу спорідненість до оксиду вуглецю (СО). У водному середовищі вільний від білкової частини гем зв'язується з СО у 25 000 разів сильніше, ніж О2. Високий ступінь спорідненості гему до СО порівняно з О2 пояснюється різним просторовим розташуванням комплексів Fe2+ гему щодо СО та О2 (рис. 5.31, А).
У комплексі Fe2+ гему з СО атоми Fe2+, вуглецю та кисню розташовані на одній прямій, а в комплексі Fe2+ гему з О2 атоми заліза й кисню розміщені під кутом, що відображає їхнє оптимальне просторове розміщення.
У міоглобіні та гемоглобіні над Fe2+ в ділянці приєднання О2 міститься Гіс Е7, що порушує оптимальне розміщення СО в центрі зв'язування білків і послаблює його взаємодію з гемом. Навпаки, той же Гіс Е7 створює оптимальні умови для зв'язування О2 (рис. 5.31, Б). У результаті спорідненість гему до СО в білках усього у 200 разів перевищує його спорідненість до О2.
Зниження спорідненості гемвмісних білків до СО має важливе біологічне значення. СО утворюється в невеликих кількостях при катаболізмі деяких речовин, у даному випадку гему.
Цей ендогенно утворений СО блокує приблизно 1 % гемвмісних білків. Якщо б спорідненість гему до СО не зменшувалась під впливом білкового оточення, ендогенний оксид вуглецю міг би викликати серйозні отруєння.
Чотири поліпептидні ланцюги, що з'єднані разом, утворюють майже правильну форму кулі, де кожен α-ланцюг контрактує з двома β-ланцюгами (рис. 5.32).
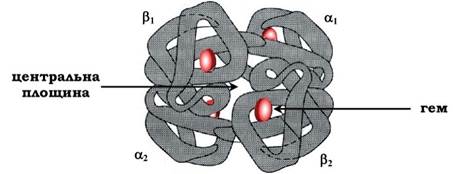
Рис. 5.32. Будова гемоглобіну
Оскільки в ділянці контакту між α1- і β1-, а також між α2- та β2-ланцюгами знаходиться багато гідрофобних радикалів, то між цими поліпептидними ланцюгами формується сильне зв'язування за рахунок виникнення в першу чергу гідрофобних, а також іонних і водневих зв'язків. У результаті утворюються димери α1β1 і α2β2. Між цими димерами в тетрамерній молекулі гемоглобіну виникають в основному полярні (іонні й водневі) зв'язки, тому при зміні рН середовища в кислий або лужний бік у першу чергу руйнуються зв'язки між димерами. Крім того, димери здатні легко переміщуватись один відносно одного.
Оскільки поверхня протомерів нерівна, поліпептидні ланцюги в центральній області не можуть щільно прилягати один до одного, у результаті в центрі формується "центральна порожнина", що проходить крізь усю молекулу гемоглобіну.
Основна функція гемоглобіну - транспорт О2 від легень до тканин. Олігомерна структура гемоглобіну забезпечує швидке насичення його киснем у легенях (утворення оксигемоглобіну - Нb(О3)4), можливість відщеплення кисню від гемоглобіну в капілярах тканин при відносно високому парціальному тиску О2, а також можливість регуляції спорідненості гемоглобіну до О2 залежно від потреб тканин у кисні.
О2 зв'язується з протомерами гемоглобіну через Fe2+, який з'єднаний із чотирма атомами азоту пірольних кілець гему та атомом азоту Гіс F8 білкової частини протомера. Зв'язування О2 з координаційним зв'язком Fe2+, що залишився вільним, відбувається на іншому боці від площини гему в області Гіс Е7 (аналогічно тому, як це відбувається в міоглобіні). Гіс Е7 не взаємодіє з О2, але забезпечує оптимальні умови для його зв'язування (рис. 5.33).
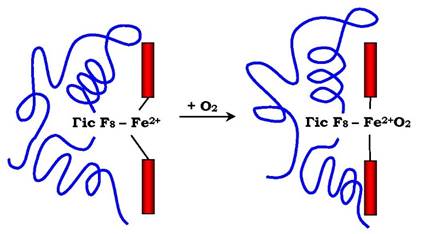
Рис. 5.33. Зміни положення Fe2+ і білкової частини гемоглобіну при приєднанні О2
У дезоксигемоглобіні завдяки ковалентному зв'язку з білковою частиною атом Fe2+ виступає з площини гему в напрямку Гіс Fe. Приєднання О2 до атома Fe2+ одного протомера викликає його переміщення в плошцну гему, за ним переміщується залишок Гіс F8 і поліпептидний ланцюг, до складу якого він входить. Оскільки протомер зв'язаний з іншими протомерами, а білки мають конформаційну лабільність, відбувається зміна конформації всього білка. Конформаційні зміни, які відбулися в інших протомерах, забезпечують приєднання наступної молекули О2, що викликає нові конформаційні зміни в білку та прискорення зв'язування наступної молекули О2. Четверта молекула О2 приєднується до гемоглобіну в 300 разів легше, ніж перша молекула (рис. 5.34).
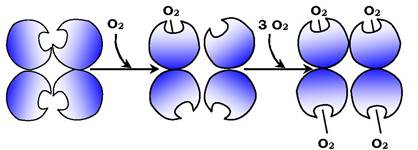
Рис. 5.34. Кооперативні зміни конформації протомерів гемоглобіну при приєднанні О2
Зміни конформації (а відповідно і функціональних властивостей) усіх протомерів олігомерного білка при приєднанні ліганду тільки до одного з них називають кооперативними змінами конформації протомерів.
Аналогічним чином у тканинах дисоціація кожної молекули О2 змінює конформацію всіх протомерів і полегшує відщеплення наступних молекул О2.
Кооперативність у роботі протомерів гемоглобіну можна спостерігати й на кривих дисоціації О2 для міоглобіну й гемоглобіну (рис. 5.35).
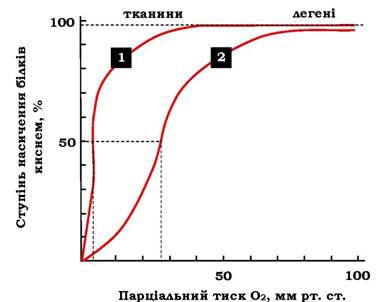
Рис. 5.35. Криві дисоціації кисню для міоглобіну й гемоглобіну залежно від парціального тиску кисню:
1 - міоглобін (Р50 = 2,8) 2 - гемоглобін (Р50 = 26)
Криві дисоціації показують, наскільки насичені дані білки О2 за різних значень парціального тиску кисню.
Крива дисоціації О2 для міоглобіну має вигляд простої гіперболи. Це вказує на те, що міоглобін зворотно зв'язується з лігандом,
1 сторонні фактори на це не впливають:
![]()
Процеси утворення та розкладу оксиміоглобіну перебувають у рівновазі, і ця рівновага зміщується вліво або вправо залежно від того, додається чи видаляється кисень із системи. Міоглобін зв'язує кисень, який у капілярах тканин звільняє гемоглобін, і сам міоглобін може звільняти О2 у відповідь на підвищення потреб у ньому м'язових тканин і при інтенсивному використанні O2 у результаті фізичного навантаження.
Міоглобін має дуже високу спорідненість до О2. Навіть при парціальному тиску О2, що дорівнює 1-2 мм рт. ст., міоглобін залишається зв'язаним з О2 на 50 %.
Із графіка на рис. 5.35 видно, що гемоглобін має набагато нижчу спорідненість до О2; напівнасичення гемоглобіну О2 настає за більших значень тиску О2 (близько 26 мм рт. ст.).
Крива дисоціації для гемоглобіну має сигмоїдну форму (S-подібну). Це вказує на те, що протомери гемоглобіну працюють кооперативно: чим більше О2 віддають протомери, тим легше відбувається відщеплення наступних молекул О2.
У капілярах м'язів у стані спокою, коли тиск О2 становить близько 40 мм рт. ст., більша частина кисню повертається у складі оксигемоглобіну назад до легень. Під час фізичної роботи тиск О2 у капілярах м'язів знижується до 10-20 мм рт. ст. Саме в такій ділянці (від 10 до 40 мм рт. ст.) розмішується "крута частина» S-подібної кривої, де найбільшою мірою виявляється властивість кооперативної роботи протомерів.
Отже, завдяки унікальній структурі, кожен із розглянутих білків пристосований виконувати свою функцію: міоглобін - приєднувати О2, що вивільнюється гемоглобіном, накопичувати його у клітині й віддавати в разі необхідності; гемоглобін - приєднувати О2 у легенях, де його насичення доходить до 100 %, і віддавати О2 у капілярах тканин залежно від зміни в них тиску О2.
Окиснення органічних речовин з метою отримання енергії відбувається в мітохондріях клітин з використанням О2, що транспортується гемоглобіном із легень. У результаті окиснення речовин утворюються кінцеві продукти розпаду СО2 і Н2О, кількість яких пропорційна інтенсивності процесів окиснення.
СО2, що утворився у тканинах, транспортується до еритроцитів. Там під дією ферменту карбангідрази відбувається збіль
шення швидкості утворення Н2СО3. Слабка вугільна кислота може дисоціювати на Н+ і HCO3 :
![]()
Рівновага реакції в еритроцитах, що містяться в капілярах тканин, зміщується вправо, оскільки протони, які утворюються в результаті дисоціації вугільної кислоти, можуть приєднуватись до специфічних ділянок молекул гемоглобіну: до радикалів Гісі4б двох β-ланцюгів, радикалів Гісі22 і кінцевих α-аміногруп двох α-ланцюгів. Усі ці шість ділянок унаслідок переходу гемоглобіну від окси- до дезоксиформи набувають більшої спорідненості до Н+ у результаті локальної зміни амінокислотного оточення навколо цих ділянок (наближення до них від'ємно заряджених карбоксильних груп амінокислот).
Приєднання трьох пар протонів до гемоглобіну зменшує його спорідненість до О2 й посилює транспорт О2 у тканини, котрі його потребують (рис. 5.36, А). Збільшення вивільнення О2 гемоглобіном залежно від концентрації Н+ називають ефектом Бора (на честь датського фізіолога Крістіана Бора, який перший відкрив цей ефект).
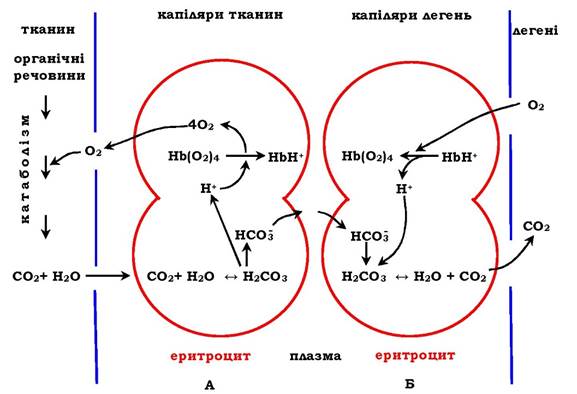
Рис. 5.36. Перенесення Н+ і СО2 з кров'ю:
А - вплив концентрації СО2 і Н+ на вивільнення О2 з комплексу з гемоглобіном у тканинах (ефект Бора);
Б - оксигенування дезоксигемоглобіну в легенях, утворення та виділення СО2
У капілярах легень високий парціальний тиск О2 приводить до оксигенування гемоглобіну й видалення шести протонів. Реакція ![]() зсувається вліво й утворений СО2 виділяється в альвеолярний простір і видаляється з повітрям, яке видихається (рис. 5.36, Б).
зсувається вліво й утворений СО2 виділяється в альвеолярний простір і видаляється з повітрям, яке видихається (рис. 5.36, Б).
Отже, молекула гемоглобіну протягом еволюції набула здатності сприймати й реагувати на інформацію, яку отримує з навколишнього середовища. Збільшення концентрації протонів у середовищі знижує спорідненість О2 до гемоглобіну й посилює його транспорт до тканин (рис. 5.37).
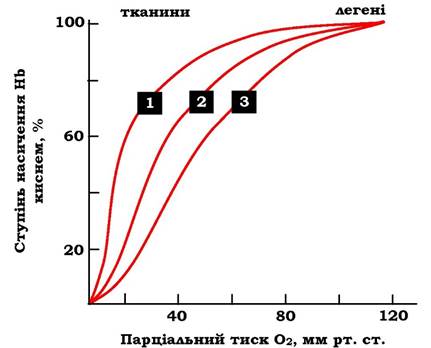
Рис. 5.37. Вплив рН на криву дисоціації О2 для гемоглобіну:
1 - рН = 7,6; 2 - рН = 7,2; 3 - рН = 6,8
Більша частина СО2 транспортується кров'ю у вигляді бікарбонату НСО3-. Невелика кількість СО2 (близько 15-20 %) може переноситися до легень, оборотно приєднуючись до неіонізованих кінцевих α-аміногруп:
![]()
у результаті утворюється карбогемоглобін, де R - поліпептидний ланцюг гемоглобіну. Приєднання СО2 до гемоглобіну також знижує його спорідненість до О2.
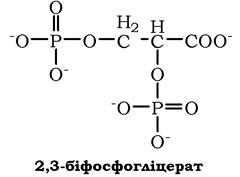
2,3-Біфосфогліцерат (БФГ) - алостеричний регулятор спорідненості гемоглобіну до О2. Ця речовина синтезується в еритроцитах із проміжного продукту окиснення глюкози 1,3-біфосфогліцерату.
У нормальних умовах 2,3-біфосфогліцерат присутній в еритроцитах приблизно в тій самій концентрації, що й гемоглобін. БФГ, приєднуючись до гемоглобіну, також може змінювати його спорідненість до О2.
У центрі тетрамерної молекули гемоглобіну є порожнина, утворена амінокислотними залишками всіх чотирьох протомерів. Центральна порожнина - місце приєднання БФГ.
Розміри центральної порожнини можуть змінюватися: відщеплення О2 від оксигемоглобіну викликає його конформаційні зміни, які сприяють утворенню додаткових іонних зв'язків між димерами α1β1 і α2β2. У результаті просторова структура дезоксигемоглобіну стає жорсткішою, напруженою, а центральна порожнина розширюється.
Поверхня порожнини обмежена залишками амінокислот, серед яких є позитивно заряджені радикали Ліз82, Гіс143β-ланцюгів і позитивно заряджені α-аміногрупи N-кінцевого валіну β-ланцюгів. БФГ, що має сильний негативний заряд, приєднується до розширеної порожнини дезоксигемоглобіну за допомогою іонних зв'язків, котрі утворюються з позитивно зарядженими функціональними групами двох β-ланцюгів гемоглобіну. Приєднання БФГ ще сильніше стабілізує жорстку структуру дезоксигемоглобіну й знижує спорідненість білка до О2 (рис. 5.38). Такий ліганд називають алостеричним, а центр, де зв'язується алостеричний ліганд, - алостеричним центром (від грец. алос - інший, інакший, стерос - просторовий).
У легенях високий парціальний тиск О2 викликає оксигенування гемоглобіну. Розрив іонних зв'язків між димерами α1β1 та α2β2приводить до "розслаблення" білкової молекули, зменшення центральної порожнини і витіснення БФГ.
![]()
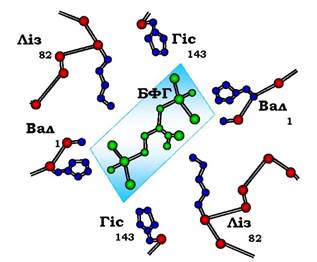
Рис. 5.38. Взаємодія 2,3-біфосфогліцерату з амінокислотними залишками центральної порожнини дезоксигемоглобіну
Концентрація БФГ в еритроцитах людей, які живуть у певних кліматичних умовах, - величина постійна. Але в період адаптації до високогір'я, коли людина піднімається на висоту понад 4000 м над рівнем моря, концентрація БФГ уже через два дні збільшується майже у два рази (від 4,5 до 7,0 ммоль/л). Це знижує спорідненість гемоглобіну до О2 і збільшує кількість О2, що транспортується до тканин (рис. 5.39).
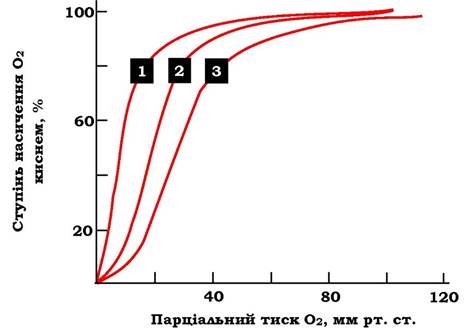
Рис. 5.39. Вплив різних концентрацій 2,3-біфосфогліцерату на спорідненість гемоглобіну до О2:
1 - гемоглобін без 2,3-БФГ; 2 - норма (2,3-БФГ = 5,0 ммоль/л); 3 - кров людини, адаптованої до висоти (2,3-БФГ - 8,0 ммоль/л)
Таку саму адаптацію спостерігають у хворих із захворюваннями легень, під час яких розвивається загальна гіпоксія тканин. Так, у хворих із важкою обструктивною емфіземою легень парціальний тиск знижується зі 100 до 50 мм рт. ст. Але при цьому в еритроцитах посилюється вироблення БФГ, і його концентрація підвищується від 4,5 до 7,0 ммоль/л, що суттєво збільшує транспортування О2 до тканин.
У крові, консервованій у деяких середовищах, наприклад цитрат-декстрозному, за десять днів концентрація БФГ знижується з 4,5 до 0,5 ммоль/л. Гемоглобін такої крові має дуже високу спорідненість до О2. Якщо кров зі зниженою концентрацією БФГ переливати важкохворим, виникає небезпека розвитку гіпоксії тканин. Уведені з кров'ю еритроцити за 24 год можуть відновити лише половину нормальної концентрації БФГ. Додаванням у кров БФГ неможливо відновити нормальну концентрацію його в еритроцитах, оскільки, маючи високий від'ємний заряд, БФГ не може проникати через мембрани еритроцитів. Тому тепер у кров додають речовини, здатні проникати через мембрану еритроцитів і підтримувати в них нормальну концентрацію БФГ.
Таким чином, олігомерний білок гемоглобін, на відміну від мо- номерного спорідненого білка міоглобіну, здатен приєднувати до специфічних ділянок чотири різні ліганди: О2, Н+, СО2 і БФГ. Усі вони приєднуються до просторово розділених ділянок, але конформаційні зміни білка в місці приєднання одного ліганду передаються на весь олігомерний білок і змінюють спорідненість до нього інших лігандів. Так, кількість О2, що надходить до тканин, залежить не тільки від парціального тиску О2, але й від концентрації алостеричних лігандів, які розширюють можливості регуляції функцій гемоглобіну.
Як розглядали вище, у капілярах працюючого м'яза збільшення концентрації СО2 і Н+ зменшує спорідненість гемоглобіну до О2 і збільшує віддачу його до тканин.
Отже, завдяки впливу регуляторних лігандів олігомерні білки здатні пристосовувати свою конформацію і функції до змін, що відбуваються в навколишньому середовищі.